Механизмы для столов раздвижные: Механизмы для раздвижных столов
Механизмы трансформации обеденных столов
Если хозяева любят отмечать праздники всей семьей или просто встречать гостей, то удобно приобрести крепкий раздвижной стол. В обычное время он будет занимать немного места, а в случае торжества размер столешницы модели можно быстро увеличить, чтобы с комфортом разместить много людей.
Стол-трансформер с цангой
Царга – соединительная деталь, которая располагается между опорами и столешницей. Она устанавливается по всему периметру и обеспечивает устойчивость конструкции. Современные технологии производства и качественная фурнитура позволяют сделать обеденный стол, который прослужит долго даже при частом раскладывании.
Мебель с прочной царгой выдерживает большие нагрузки как в собранном, так и разобранном состоянии. Если еще используется металлическая фурнитура, то изделие может эксплуатироваться несколько десятков лет. Такой стол выглядит солидно и массивно, что позволяет ему гармонично вписаться в классический интерьер. Но для некоторых дизайнерских задумок излишняя тяжеловесность модели окажется нежелательной. При выборе стола стоит обратить особое внимание на удобство царги. Если она сконструирована плохо, то обедающим будет неудобно закидывать ногу на ногу.
Но для некоторых дизайнерских задумок излишняя тяжеловесность модели окажется нежелательной. При выборе стола стоит обратить особое внимание на удобство царги. Если она сконструирована плохо, то обедающим будет неудобно закидывать ногу на ногу.
Существует несколько типов исполнения раздвижной столешницы. В зависимости от них изделия будут отличаться по простоте раскладывания, площади дополнительных поверхностей, а также внешнему виду.
Индивидуальная центральная вставка
Это надежная, долговечная конструкция. В сложенном виде у мебели посередине столешницы есть стыковочный шов, а в разложенном – их становится два. Если не присматриваться к такой мелочи, то у стола не будет визуальных недостатков.
Принцип устройства раздвижного механизма довольно прост. Вставка располагается в незакрепленном состоянии под столешницей, в центре. Когда стол надо разложить, боковины поверхности раздвигаются, а центральная часть вынимается и кладется в специальные пазы.
Подобные столы, изготовленные много лет назад, до сих пор служат своим хозяевам, так как сломать их довольно непросто. Но в процессе эксплуатации у них появляются дефекты – боковые части, двигающиеся по деревянным пазам, могут переклинить или перекоситься. Поэтому в современных изделиях с центральной вставкой устанавливается металлические направляющие. Это не только решает проблемы с заклиниванием боковин, но и обеспечивает легкость хода при раскладывании и складывании изделия.
Вставка по центру с поворотным механизмом
У такого стола центральную вставку делят на две части, которые могут поворачиваться. Когда изделие собрано, оно выглядит так же, как и с обычной вставной частью. Когда модель раскладывается, мест стыковки становится больше. Механизм раздвижения подобен описанному выше.
Преимущество такого поворотного механизма по сравнению с предыдущей моделью – высокая скорость разбора стола. Современная фурнитура и технологии изготовления позволяют хозяевам с легкостью разложить стол, потратив на это совсем немного времени – не больше одной минуты.
Двойная столешница
Особенность такого изделия – простота механических компонентов. В сложенном состоянии столешница смотрится несколько громоздко, так как ее толщина увеличена вдвое. Чтобы ее разложить, достаточно приподнять верх, а второй слой поверхности просто потянуть в сторону. Центральная часть столешницы уложится в свободное пространство и будет готова к использованию. Площадь поверхности у такого стола увеличивается ровно в два раза.
К недостаткам такой конструкции можно отнести невысокую прочность. Стол нельзя подвергать серьезным нагрузкам, так как он может этого не выдержать и сломаться. Возможно, именно поэтому такой вариант не является распространенным.
Стол нельзя подвергать серьезным нагрузкам, так как он может этого не выдержать и сломаться. Возможно, именно поэтому такой вариант не является распространенным.
Раздвижная модель с перемещением опор
У такого стола принцип увеличения площади столешницы принципиально отличается. Если у перечисленных вариантов двигалась только столешница, то теперь у изделия будут перемещаться и ножки. Для раскладывания нужно взяться снизу за царгу и раздвинуть ее. Добавочная часть у столешницы может или поворачиваться, или раздвигаться. Площадь модели увеличивается от 30 до 50%. Изделие раздвигается легко и характеризуется устойчивостью. Но чтобы оно выглядело эстетично в разложенном состоянии, рабочую поверхность лучше декорировать скатертью.
Раскладная модель с перемещением опор
У такого стола тоже двигаются опоры. В сложенном виде изделие выглядит так же, как и модель с двойной столешницей. Для раскладывания царгу тоже необходимо раздвинуть. Полностью передвинув опоры, надо раскрыть столешницу по принципу «книжки» и поместить ее по центру. Это намного удобнее делать, если у конструкции есть подъемное механическое устройство. Плюсом подобного стола является существенное увеличение площади (в два раза), а минусом – невысокая прочность. Изделие ни в коем случае нельзя перегружать в разложенном виде.
Полностью передвинув опоры, надо раскрыть столешницу по принципу «книжки» и поместить ее по центру. Это намного удобнее делать, если у конструкции есть подъемное механическое устройство. Плюсом подобного стола является существенное увеличение площади (в два раза), а минусом – невысокая прочность. Изделие ни в коем случае нельзя перегружать в разложенном виде.
Стол-трансформер без царги
У этого типа столов нет массивного основания под рабочей поверхностью, поэтому они смотрятся грациозно и стильно и превосходно вписываются в популярные интерьерные стили. Часто для подобных моделей столешница делается из прозрачных материалов.
Стеклянная мебель
Такие модели – современны и эстетичны, они подходят для разных дизайнерских решений. Раскладывание изделия происходит посредством металлических направляющих, спрятанных под столешницей. Двигаются элементы очень легко – для их смещения достаточно слегка потянуть поверхность. Раздвижные столы из стекла не только радуют глаз, но и отличаются практичностью, так как площадь их столешницы может увеличиться до двух раз. Если поверхность становится намного больше, то модель выпускают с дополнительными опорами для большей устойчивости.
Если поверхность становится намного больше, то модель выпускают с дополнительными опорами для большей устойчивости.
Недостатком стеклянных изделий считается невысокий запас прочности: и направляющие детали, и столешница не приспособлены к выдерживанию больших нагрузок. Также многих покупателей смущает существенная стоимость мебели – качественные комплектующие части, обычно произведенные на импортном оборудовании или за рубежом, просто не могут стоить дешево.
Стол-книжка
Еще можно отметить стол-трансформер, своей формой напоминающий книжку. В сложенном виде его можно использовать в качестве журнального столика или тумбы. А в разложенном состоянии модель превращается в удобный обеденный стол.
Как выбрать подходящий стол-трансформер
Прежде чем совершить покупку, следует:
- Измерить размеры помещения. Стоит обратить внимание на то, сколько пространства займет стол в сложенном виде и насколько гармонично он будет выглядеть в интерьере.
 Если правильно выбрать габариты, модель будет использоваться постоянно в собранном виде и удобно раскладываться в праздники при приходе гостей. А вот переплачивать за огромную столешницу, которая использоваться не будет, не стоит.
Если правильно выбрать габариты, модель будет использоваться постоянно в собранном виде и удобно раскладываться в праздники при приходе гостей. А вот переплачивать за огромную столешницу, которая использоваться не будет, не стоит.
 Если правильно выбрать габариты, модель будет использоваться постоянно в собранном виде и удобно раскладываться в праздники при приходе гостей. А вот переплачивать за огромную столешницу, которая использоваться не будет, не стоит.
Если правильно выбрать габариты, модель будет использоваться постоянно в собранном виде и удобно раскладываться в праздники при приходе гостей. А вот переплачивать за огромную столешницу, которая использоваться не будет, не стоит.- Оценить функциональность изделия. Надо обдумать, хватит ли увеличенной площади столешницы для приходящих родственников и друзей. Можно посмотреть, есть ли какие-то удобные дополнительные возможности – выдвижные ящики для хранения столовых приборов, небольшие полки для салфеток и мелочей.
- Проверить качество материалов и фурнитуры, из которых изготовлен стол. Если комплектующие ненадежны, конструкция быстро сломается. Поэтому не нужно гнаться за дешевизной и отдавать деньги за недолговечные изделия.
Чем серьезнее обдумать покупку раздвижного обеденного стола, тем дольше он прослужит, порадует хозяев и гостей и даст возможность приятно провести время за зваными обедами и застольными беседами.
механизмы, материалы, стили. Салоны ТРИО-Интерьер в Москве.
09 Августа 2017 3386
<p>
Если вы решили купить <a href=»/tags/razdvizhnye_stoly/»>раздвижной стол</a>, чтобы принимать гостей красиво и при этом сэкономить место в квартире или загородном доме, у вас есть возможность выбора.
</p>
<p>
Как его сделать правильно и по каким критериям оценивать столы – об этом расскажут наши дизайнеры и технические специалисты.
<h3>Виды раздвижных столов</h3>
<p>
Начнем с практической функции раздвижных столов, ведь в первую очередь важна их способность менять размер.
 Итак, принцип тот же – <a href=»/tags/malenkie_obedennye_stoly_/»>маленький стол</a> превращается в большой. В собранном виде он великолепно экономит место на кухне, гостиной или в столовой, в разложенном дает возможность усадить с комфортом до 10-14 человек. Если вы экономите место или у вас не так часто бывают гости, раздвижной стол, — то, что нужно.
Итак, принцип тот же – <a href=»/tags/malenkie_obedennye_stoly_/»>маленький стол</a> превращается в большой. В собранном виде он великолепно экономит место на кухне, гостиной или в столовой, в разложенном дает возможность усадить с комфортом до 10-14 человек. Если вы экономите место или у вас не так часто бывают гости, раздвижной стол, — то, что нужно.</p>
<p>
Как правило, обеденные столы стандартной высоты увеличиваются за счет выдвигающихся «лепестков», которые хранятся под столешницей или в ней – они легко выезжают из-под нее и благодаря плотной стыковке обеспечивают увеличение стола на запланированную длину. При этом стол может менять свою форму – например, с квадратной на овальную как <a href=»/catalog/stolovye_gruppy/obedennye_stoly/23433/»>Titan III 1136</a> (Draenert). <br>
<p>
<a href=»/catalog/stolovye_gruppy/obedennye_stoly/23433/»><img alt=»Стол Titan III 1136 от Draenert» src=»https://www.
 trio.ru/upload/iblock/387/38734b74ddbded27a8648968c6f6eed0.jpg» title=»Titan III 1136″></a><br>
trio.ru/upload/iblock/387/38734b74ddbded27a8648968c6f6eed0.jpg» title=»Titan III 1136″></a><br></p>
<p>
Зачастую дополнительные элементы (вытяжные вкладки), увеличивающие поверхность стола, хранятся в самой столешнице, как в модели <a href=»/catalog/stolovye_gruppy/obedennye_stoly/17189/»>8810</a> (Rolf-Benz). Огромный плюс — максимум свободного пространства для ног. <br>
</p>
<p>
<a href=»/catalog/stolovye_gruppy/obedennye_stoly/17189/»><img alt=»Стол 8810 от Rolf-Benz» src=»https://www.trio.ru/upload/iblock/627/7d6923a54bb39da22316aea19bc2c9cf.jpg» title=»8810″></a><br>
</p>
<p>
Иногда поверхность стола увеличивается за счет подъемной части – из вертикального положения она переводится в горизонтальное и закрепляется как в модели <a href=»/catalog/stolovye_gruppy/obedennye_stoly/12691/»>DAYTONA</a> (Cattelan). <br>
</p>
<p>
<a href=»/catalog/stolovye_gruppy/obedennye_stoly/12691/»><img alt=»Стол Daytona от Cattelan» src=»/upload/medialibrary/fd8/fd815d61c8dbe681190265659e5f3c4e.
 jpg» title=»Daytona»></a><br>
jpg» title=»Daytona»></a><br></p>
<h3>10 важных особенностей современных раздвижных столов</h3>
<p>
Если вспомнить советские образцы, то в противовес их функциональности можно было поставить вес стола, сложную транспортировку по квартире, не очень удобные и ненадежные механизмы раскладывания, неэкологичные и недолговечные материалы. Что же отличает современные дизайнерские столы?
</p>
<h4>Эстетика</h4>
<p>
Они действительно красивые и будут служить украшением интерьера, куда бы их ни поставили – на кухню, в столовую, гостиную, спальню и т.д.
</p>
<h4>Интуитивно понятный механизм раскладывания.</h4>
<p>
Вам не придется долго изучать инструкцию, чтобы понять, как обращаться со столом.
</p>
<iframe src=»https://www.youtube.com/embed/R9dGaBdJX1c» frameborder=»0″ allowfullscreen></iframe>
<h4>Легкость в использовании</h4>
<p>
Даже сложные механизмы не требуют большой физической силы.
 С любым, даже очень тяжелым столом, справится и хрупкая женщина, и в этом большой плюс.
С любым, даже очень тяжелым столом, справится и хрупкая женщина, и в этом большой плюс.</p>
<h4>Значительное увеличение в размере</h4>
<p>
В некоторых моделях стол может увеличиться до 4 метров. По ссылке — <a href=»/tags/bolshie_obedennye_stoly/»>большие обеденные столы</a>.
</p>
<hr color=»00a999″ size=»3″>
</br>
<p align=»center»>У нас вы можете подобрать обеденный стол любого дизайна и стиля из 600 наименований!</p>
<p align=»center»><a href=»/catalog/stolovye_gruppy/obedennye_stoly» target=»_blank»>Выбрать стол</a></p>
<hr color=»00a999″ size=»3″>
</br>
<h4>Регулируемая основа</h4>
<p>
Также частая опция в современных моделях. Вы можете выставить нужную высоту стола и поменять ее при необходимости.
</p>
<h4>Высокая степень комфорта</h4>
<p>
Эргономика современных столов продумана до мелочей, поэтому сидеть за ними удобно.
 Что немаловажно – какой бы механизм раскладывания не предложил производитель, всегда будет достаточно места для ног.
Что немаловажно – какой бы механизм раскладывания не предложил производитель, всегда будет достаточно места для ног.</p>
<h4>Разнообразие материалов</h4>
<p>
Современные производители используют даже сложные и, казалось бы, нетипичные для столов материалы – <a href=»/tags/kamennye_stoly/»>камень</a>, стекло, керамику и т.д. <br>
</p>
<p>
<img alt=»Пример раскладывания стола Draenert» src=»/upload/medialibrary/0b2/0b25ed0196eb06a2b022c554e2cafe3d.gif» title=»Раскладной стол Draenert»><br>
</p>
<h4>Разнообразие цветовых решений</h4>
<p>
Ваша столешница может быть любого цвета и идеально вписываться в интерьер.
</p>
<h4>Легкость перемещения</h4>
<p>
Некоторые из современных раскладывающихся столов оснащены колесиками, поэтому легко перемещаются в пространстве.
</p>
<h4>Срок эксплуатации до 15-20 лет</h4>
<p>
Обеспечивается современными материалами и технологиями.

</p>
<p>
<i><b>Важно!</b></i> Среди наиболее востребованных моделей – раздвижные <a href=»/tags/obedennye_steklyannye_stoly/»>столы со стеклянной столешницей</a>. Вопреки нашим представлениям о хрупкости стекла и необходимости крайне осторожно обращаться с этим материалом, такой стол выдерживает значительные нагрузки и долго остается в великолепном состоянии. Однако имейте в виду, что на ощупь стекло будет холодным. Обратите внимание на <a href=»/catalog/stolovye_gruppy/obedennye_stoly/14961/»>Ray plus</a> (Fiam, Италия). <br>
</p>
<p>
<a href=»/catalog/stolovye_gruppy/obedennye_stoly/14961/»><img alt=»Стол Ray plus от Fiam» src=»/upload/medialibrary/e3c/e3c41321341a5e46a6a7471d330fac0a.jpg» title=»Ray plus»></a><br>
</p>
<h3>Раскладывающиеся столы в интерьере</h3>
<p>
Благодаря новомодным техническим решениям у потребителя есть превосходный выбор.
 Вы выбираете не только механизм раскладывая и финальную длину стола, но также материал и цвет. Стоит отметить, что раздвижные столы нового поколения отлично смотрятся и в классических, и в современных интерьерах. Востребовано сочетание различных материалов столешницы – камня и керамики, <a href=»/tags/stoly_iz_stekla_i_dereva/»>дерева и стекла</a>, и т.д.
Вы выбираете не только механизм раскладывая и финальную длину стола, но также материал и цвет. Стоит отметить, что раздвижные столы нового поколения отлично смотрятся и в классических, и в современных интерьерах. Востребовано сочетание различных материалов столешницы – камня и керамики, <a href=»/tags/stoly_iz_stekla_i_dereva/»>дерева и стекла</a>, и т.д.</p>
<p>
<i><b>Совет:</b></i> Чтобы точнее определиться с выбором, задайте себе вопрос – какие тактильные ощущения вам хотелось бы получать, ведь обедая за этим столом, вы все время будете дотрагиваться до столешницы. Сложность ухода за материалом также важна — раскладной стол такого качества будет служить вам на протяжении десятилетий.
</p>
<p>
Исключительный пример – столы <a href=»/brands/draenert/»>Drаenert</a>. Этот производитель из Германии специализируется на современных столах с каменной столешницей, оснащенных механизмом раскладывания.
 Основа – полированная высококачественная сталь, камень – натуральный мрамор с различной обработкой поверхности. Вес такой мебели – около 300 килограмм. Такие <a href=»/tags/mramornye_stoly/»>мраморные столы</a> великолепно подчеркивают статус интерьера, создают ощущение спокойствия и надежности, легко перемещаются в пространстве квартиры или дома благодаря выдвижным колесикам. Механизм их раскладывания очень прост и не требует серьезных физических усилий.
Основа – полированная высококачественная сталь, камень – натуральный мрамор с различной обработкой поверхности. Вес такой мебели – около 300 килограмм. Такие <a href=»/tags/mramornye_stoly/»>мраморные столы</a> великолепно подчеркивают статус интерьера, создают ощущение спокойствия и надежности, легко перемещаются в пространстве квартиры или дома благодаря выдвижным колесикам. Механизм их раскладывания очень прост и не требует серьезных физических усилий.</p>
<iframe src=»https://www.youtube.com/embed/y8rciice9lU» frameborder=»0″ allowfullscreen></iframe>
<p>
<b>Консультанты салонов ТРИО знают все про столы для ваших гостиных. Приезжайте к нам, трогайте, раздвигайте и выбирайте!</b>
</p>
Виды раскладных столов и основные правила их выбора
Механизмов раскладки стола множество. По принципу действия их можно классифицировать на две базовые группы:
1. Ручные системы. Для увеличения стола нужно выполнить вручную несколько манипуляций:
- отодвинуть столешницу;
- вынуть (взять с места хранения) дополнительную вставку;
- расположить в предусмотренном для нее месте;
- зафиксировать.

Складывание стола выполняется в обратном порядке.
Ручным механизмом может снабжаться и столешница без вставки. Она представляет собой конструкцию из двух модулей, соединенных рояльными, карточными или секретерными петлями и положенных друг на друга. При необходимости увеличить размеры крышки ее поворачивают и распахивают.
Ручными механизмами оснащены практически все раскладные столы экономкласса.
| Рисунок 2 — Раскладывание стола с ручным механизмом без вставки | Рисунок 1 — Раскладывание стола с ручным механизмом и вставкой |
2. Полуавтоматические системы. Чтобы раскрыть или сложить столешницу, достаточно потянуть за специальную ручку или нажать на деталь в определенном месте конструкции, — механизм выполнит работу самостоятельно. Такой мебелью удобнее пользоваться, но она значительно дороже. Обычно узлами автоматики снабжены лишь итальянские столы премиум-класса или люксовая мебель, выполняемая на заказ в дизайнерских мастерских.
Разновидности ручных и полуавтоматических систем увеличения столешниц
Теперь рассмотрим максимально популярные и практичные разновидности раскладки, которые могут использоваться и для ручных, и для полуавтоматических механизмов.
Центральная вставка. Дополнительный модуль (или несколько) вставляется в центр столешницы, половины которой раздвигаются в противоположные стороны. Для компактного хранения вставки предусмотрена ниша, где деталь находится, когда стол сложен. Преимущества конструкции — сравнительная дешевизна, простота использования. Стол с центральной вставкой удобен для размещения в центре комнаты или у стены.
Вставка-бабочка — вариация центральной. Жестко зафиксированная к столу дополнительная деталь складывается пополам. Это удобнее, чем полностью съемная вставка, и практично, если стол периодически приходится перемещать: вставка-бабочка не выпадает, не шатается при движении. Благодаря этому исключен риск повреждения мебели при транспортировке.
| Модель Tivoli, столешница с центральной вставкой-«бабочкой» | Стол Эссен, с боковым крылом, и выдвигающимися ногами | Стол Сидней, с одной центральной вставкой |
Стол-консоль. Интересное конструктивное решение: раздвигается не только столешница, но и рама, за счет чего сохраняется устойчивость и прочность. У консолей может быть до 7 вставок. Минус — необходимость хранения съемных деталей отдельно от стола.
Боковое крыло. Конструкция предусматривает 1 или 2 дополнительных элемента, соответственно, приставленных с одной либо с двух сторон основной столешницы. Она при одном крыле может сдвигаться, при двух — быть стационарной. Преимущества механизма — простота, экономичность.
Есть модели столов с боковым крылом, в которых смещается не столешница, а пара ножек, выдвигаясь вместе с рамой вбок. Вставки располагаются на получившемся удлинении конструкции.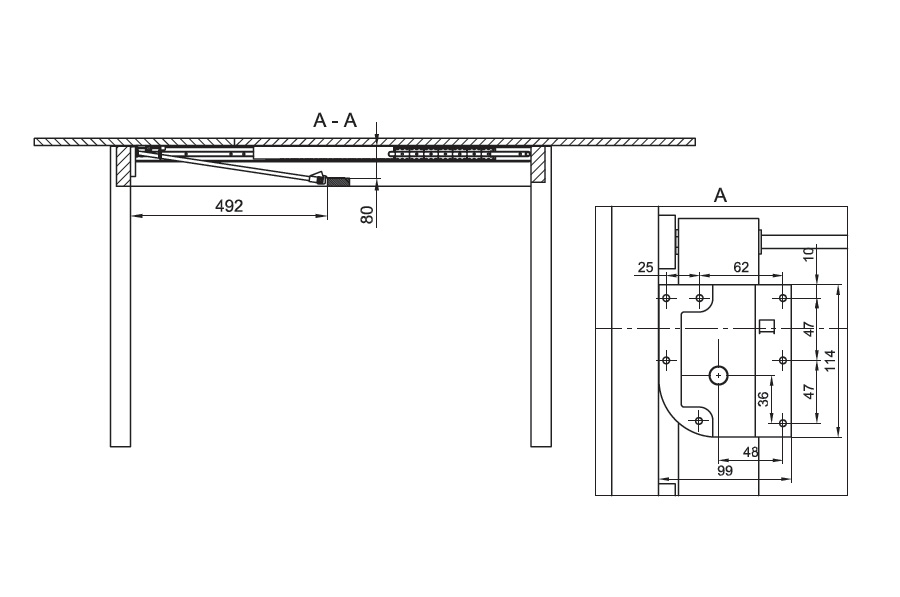 Возможно увеличение одного или обоих краев. Удобно, что ножки стола всегда остаются по бокам. Минус — такой механизм удорожает мебель.
Возможно увеличение одного или обоих краев. Удобно, что ножки стола всегда остаются по бокам. Минус — такой механизм удорожает мебель.
Стол-слайдер. Здесь столешница тоже увеличивается за счет бокового элемента, но его размер равен или почти равен габаритам основной детали. Когда стол сложен, дополнительный модуль находится под крышкой, при раскладывании — выезжает и поднимается на ее уровень. Плюс механизма — простота использования, недостаток — дороговизна производства, повышающая цену мебели.
| Модель Poket со слайдер-механизмом | Модель NEW Mistery: 3 вставки, каждая увеличивает стол-консоль на 50 см |
Стол-книжка. Механизм так назван благодаря сходству элементов столешницы со страницами. Крышка состоит из соединенных петлями двух частей, в сложенном состоянии находящихся друг над другом. При раскладке они как бы распахиваются, увеличивая поверхность в 2 раза.
Консоль-книжка — вариация стола-книжки. У консоли обычно не одно, а два крыла, они значительно больше, прикреплены петлями к длинным сторонам основной столешницы. Поставленная у стены консоль-книжка экономит место в комнате, но для полного раскрывания мебель приходится отодвигать, поэтому для удобства она часто снабжается колесиками.
Модель «Гавана»: 2 крыла консоли в развернутом состоянии образуют овальную столешницу
| Консоль-книжка «Гавана», с двумя боковыми крыльями | Консоль-книжка Mascotte, с изменяемой высотой и столешницей-книжкой |
Сателитный механизм. Используется редко, в основном в итальянских столах премиум-класса. В конструкции предусмотрена пара боковых крыльев, которые при раскладывании выдвигаются из-под основной детали по кругу, для этого необходимо много свободного пространства. Плюсы — оригинальный вид, простота трансформации. Минус — высокая цена.
Минус — высокая цена.
2 главных правила выбора раскладного стола
Даже раскладной стол может оказаться громоздким и неудобным, если перед покупкой не продумать важные моменты.
1. Пропорциональность. Сопоставьте габариты сложенного и раскрытого стола с предназначенной для него площадью. Предварительные расчеты позволят определить, вмещается ли мебель в отведенное пространство.
Важно: от раскрытой столешницы до стен и стационарной мебели должно оставаться не менее 80 см, чтобы гости могли свободно отодвигать стулья, садясь за стол или выходя из-за него.
2. Частота пользования. Если в квартире нет гостиной или столовой и нужно экономить пространство, самым компактным решением станет консоль — перед приемом гостей ее можно выдвинуть и разложить, в остальное время она будет стоять у стены. При наличии свободной площади лучше купить большой стол, выбирая механизм раскладывания по личным предпочтениям.
При наличии свободной площади лучше купить большой стол, выбирая механизм раскладывания по личным предпочтениям.
Виды раздвижных и раскладных столов
Стол – центральный объект кухни и нередко гостиной. Это место сбора всей семьи за трапезой, праздниками, обсуждением новостей… а в доме гостеприимных хозяев это изделие — атрибут №1. Бытует выражение, что желанного и уважаемого гостя хотят видеть во главе исключительно стола, а не какой-либо иной мебели. Поэтому главными героями нашего обзора станут именно столы. Но не простые, а столы-трансформеры – раздвижные и раскладные.
Элегантная компактность и внушительная вместимость
Почему мы решили рассмотреть именно раздвижные и раскладные столы? Все довольно просто объясняется: в сложенном, как и в разложенном состоянии изделие занимает ровно столько место, сколько нужно в данный момент времени. Звучит немного фантастически и запутанно, не так ли? Но достаточно взглянуть на картинку ниже, и все становится на свои места.
И раздвижной, и раскладной стол относятся к разряду мебельных изделий-трансформеров лишь с разницей в способе собственно трансформации и механизмами для раздвижных столов: в первом случае столешницу нужно раздвинуть для увеличения полезной площади поверхности, во втором – поднять и зафиксировать боковые створки.
Поколение, чье детство прошло во времена СССР или вскоре после его развала, наверняка помнит советские раскладные столы-бабочки или древние, массивные раздвижные столы. Они не отличались эргономичными функциональными механизмами и дизайнерскими излишками.
Рассмотрим же раздвижные и раскладные столы, представленные сегодня на мебельном рынке, ознакомимся с их конструктивными особенностями. Не обойдем стороной также преимущества и недостатки каждой разновидности.
Раздвижной стол с царговым каркасом – классическое мебельное решение
Царговый каркас – конструкция, история которой берет свое начало в Европе эпохи Возрождения. Это массивное основание на ножках, поверх которого устанавливается столешница с дополнительными механизмами увеличения площади последней.
Это массивное основание на ножках, поверх которого устанавливается столешница с дополнительными механизмами увеличения площади последней.
К преимуществам безусловно относится надежность, устойчивость и долговечность. За несколько веков подобный конструктивный тип эволюционировал и модернизировался; изменялись и совершенствовались раздвижные функциональные механизмы.
Но царговые столы не лишены недостатков: из-за своих габаритов они массивно выглядят, что делает их присутствие неуместным в помещениях с преобладающим «парящим» дизайном. Также некоторые модели могут стать неудобными для высоких людей: при желании запрокинуть ногу они могут испытать некоторый дискомфорт и неудобство.
Безцарговый стол
Приемник раздвижных столов с царговым каркасом. Это порождение современной инженерной и дизайнерской мысли: для того, чтобы увеличить полезную площадь, нет необходимости в массивном основании. Но в качестве материалов для столешницы используются легкие материалы – стекло или ДСП.
Выглядит конструкция действительно яркой и парящей, легко разбирается и собирается. Но надежность ее, по сравнению с массивным царговым каркасом, проигрывает на несколько позиций. Также стоимость подобных изделий значительно выше. На ценовую политику влияют материалы каркаса и столешницы, а также облегченная функциональная фурнитура.
Раздвижные столы с отдельными и поворотными центральными вставками
Обе разновидности – не новинка на мебельном рынке. Прочная, устойчивая конструкция раздвижного стола с отдельной центральной вставкой закономерно эволюционировала в вставку поворотную, что показано на изображениях ниже.
В первом случае дополнительная секция представляет собой монолитную вставку, и хранится под столешницей, когда не используется. Чтобы увеличить стол, достаточно раздвинуть створки, вставить дополнительную центральную в специальные пазы.
В втором – дополнительная секция – это та же центральная ставка, но разделенная на две симметричные половины снабжена фурнитурой, сводящей к минимуму прилагаемые физические усилия и исключающей перекос, который нередко возникал у предшественников. Но в то же время, на поверхности появился дополнительный стыковочный шов, что может прийтись не по душе перфекционистам и педантам.
Но в то же время, на поверхности появился дополнительный стыковочный шов, что может прийтись не по душе перфекционистам и педантам.
Раздвижной стол с перемещаемыми ножками
Простая и эргономичная конструкция. Площадь столешницы увеличивается за счет специальных направляющих без приложения силы. Несмотря на функциональность, несколько страдает эстетика: заметны переходы между рамами, выдвижная столешница несимметрична основной.
Раскладной стол с перемещаемыми ножками и стол-книжка
Обе разновидности легко маскируются под журнальный столик и в случае необходимости увеличивают полезную площадь столешницы в 2 и даже более раза.
И это далеко не все разновидности. Но какую бы вы не выбрали, главное, чтобы ваш выбор приносил вам исключительно теплые и позитивные эмоции!
Как устроены столы-трансформеры, Москва
Эксперты нашего интернет-магазина подготовили для вас обзор, как выбрать самый подходящий трансформируемый журнальный или обеденный стол для своего дома.
Стол-трансформер по ГОСТ
В обиходе под столами-трансформерами понимают все изделия, способные изменять свои размеры (площадь столешницы), форму и высоту. С этим мнением согласны и ГОСТ 20400-80 и ТР ТС 025/2012, которые гласят, что «Трансформируемая мебель — мебель, конструкция которой позволяет путем перемещения деталей (элементов), менять ее функциональное назначение и (или) размеры». Под функциональностью подразумевается назначение, т.е. обеденный, кофейный, туалетный стол и т.д.
Однако в профессиональной среде сложилось более узкое деление. Под столами-трансформерами понимают, как правило, столы, которые могут изменять и высоту, и размеры столешницы. А изделия, у которых изменяется исключительно площадь столешницы, относят к раскладным столам.
В этой статье мы рассмотрим оба вида столов-трансформеров.
Стол-трансформер в обиходном значении — меняет свою функциональность
Стол Д.Рентгена, XVIII век
Стол-трансформер в узкопрофессиональной среде — меняет размах столешницы и высоту
Стол B2109-3 фабрики Gomlden
Виды столов-трансформеров
Разнообразие столов-трансформеров обусловлено широким спектром их предназначения. Для выбора подходящего трансформера, в первую очередь, ответьте на вопрос «какие два стола нужно объединить в один»: журнальный с обеденным, кухонный с гостиным, пристенный с застольным? Все зависит от назначения будущего стола, от того, какую роль он будет играть постоянно, а какую — время от времени. Варианты могут быть такими:
Для выбора подходящего трансформера, в первую очередь, ответьте на вопрос «какие два стола нужно объединить в один»: журнальный с обеденным, кухонный с гостиным, пристенный с застольным? Все зависит от назначения будущего стола, от того, какую роль он будет играть постоянно, а какую — время от времени. Варианты могут быть такими:
- журнально-обеденные столы-трансформеры,
- обеденные столы-трансформеры,
- столы-консоли,
- столы-книжки.
Во вторую очередь, обозначьте требования по занимаемой площади: где будет стоять стол, сколько места ему будет выделено, нужно ли будет прятать в шкаф или он сможет быть полноправным предметом интерьера. Исходя из этих требований подберите подходящие варианты в каталоге магазина (см.следующую иллюстрацию):
В полученную выборку могут попасть как столы-консоли, так и раскладные кухонные столы — всё зависит от заданных размеров.
В третью очередь, чтобы отсеять столы, не подходящие по сложности трансформации, задайте допустимые параметры:
- кто будет его «разворачивать» и «сворачивать», хватит ли физических сил и сноровки, ведь столы-трансформеры могут весить до 70 кг;
- насколько высоки требования к эстетике — удовлетворит ли «простенький» вариант или крайне важен абсолютно безупречный внешний вид как в собранном, так и в разобранном состоянии.

О том, как механизмы трансформации влияют на удобство раскладки и совершенство внешнего вида столов, читайте ниже, а сейчас пробежимся по их разновидностям.
Журнальные и журнально-обеденные столы
Деление столов-трансформеров на журнальные и журнально-обеденные условно, поскольку все они выполняют одинаковую задачу. К ним условно можно «присоединить» чайные и кофейные столики (из-за их небольшого размера). Впрочем, пару различий можно найти:
- чайные и кофейные столики выше журнальных;
- в журнальных принято предусматривать под столешницей места хранения для периодики (журналов и газет).
В целом же все перечисленные типы столов-трансформеров составляют очень широкую и весьма разнообразную линейку. В зависимости от конструктивных особенностей, они способны изменять площадь столешницы, ее высоту, а в некоторых случаях — и конфигурацию, т.е. взаимное расположение элементов в объеме пространства.
Журнально-обеденный трансформер может быть обеденным
Стол B2442-3 от ESF
Журнальный трансформер остаётся журнальным
Столик Агат 24. 10 фабрики ДаВита
Обеденные столы-трансформеры
Именно в этой категории, пожалуй, стоит искать универсальный стол-трансформер, который годится для гостиной любого размера, включая малогабаритное жилье. Особенно интересны при добавочных требованиях варианты с «закрытыми» местами для хранения. Обеденные столы-трансформеры — это удивительно разнообразная по стилевым решениям и выбору материалов группа. В ней можно найти все, начиная от классики и заканчивая решениями в стиле хай-тек или модерн. То есть, в зависимости от необходимости, подобрать основу для столовой группы или вариант самостоятельного предмета интерьера.
Обеденный стол-трансформер может достигать 2 метров в длину
Стол Optimata 327 Florida
Стол-консоль
Стол-консоль — изумительно универсальный предмет мебели. Его можно органично вписать в абсолютно любое помещение, включая прихожую, и он везде будет к месту. Причем при изначально миниатюрных габаритах в разложенном состоянии консоль способна принять поистине внушительные размеры.
Стол-консоль раскладывается от 44 см до 229 см в длину
Стол В2307-1 от Gomlden
Столы-книжки
Стол-книжка (или стол-тумба) безусловно хороший вариант для малогабаритной квартиры. Не такой эффектный, как консоль, зато практичный. Особенно если в нем предусмотрены места для хранения, как в изделии на фото ниже. Согласитесь, почти у каждого из нас есть сервиз — вроде бы он нужен, но занимает так много места, которого и так вечно не хватает. Конечно, место такому столу — в кухне.
И именно среди столов-книжек можно найти потрясающе компактные изделия, которые в сложенном состоянии занимают места чуть ли не как гладильная доска, то есть их можно поставить даже в крохотную кладовую и вытаскивать лишь по мере необходимости.
Стол-книжка с ящиками для хранения
Ультратонкий стол-книжка
Как устроены столы-трансформеры?
Здесь мы рассмотрим, как устроены столы-трансформеры, чем отличаются друг от друга, и какой из них лучше выбрать по параметрам, обозначенным в начале статьи.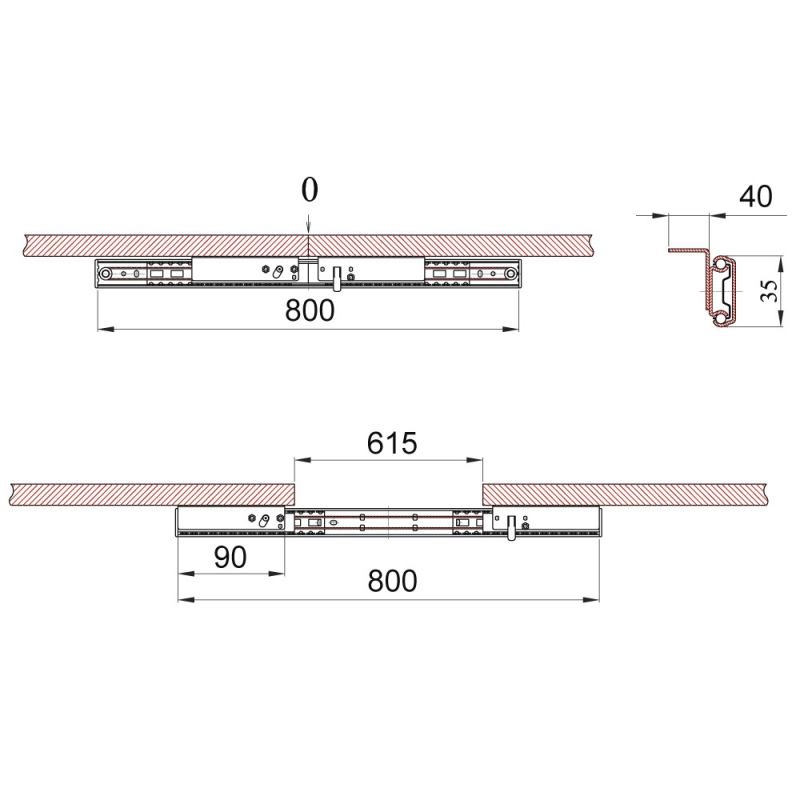
Оглавление раздела:
- Механизмы трансформации столешницы — общий обзор.
- Вкладки для столешниц и их различия.
- Раздвижные механизмы.
- Раскладные механизмы.
- Механизмы подъёма столешницы.
Механизмы трансформации столешницы
Именно от механизмов трансформации зависит удобство и простота раскладывания/складывания стола. Их можно разделить на механизмы трансформации столешницы и механизмы ее подъема. Перечислим основные элементы столов, используемые в данной статье:
- столешница — цельная или разрезная горизонтальная верхняя плита;
- полустолешница — половинка столешницы;
- вставка — «нестационарное» дополнительное полотно, используемое для увеличения площади столешницы;
- царга — вертикальный «пояс», служащий для соединения и крепления опор в царговых столах;
- подстолье — опоры и рама (или крестовины и пр.), на которые укладывается столешница;
- уголок, ролик или пандус для хранения вставок;
- опоры — ножки или боковые вертикальные панели.

Вкладки для столешниц
Цельная вкладка — съемный элемент столешницы, выполненный из единого «листа». Чаще это центральная вкладка, также одним из вариантов можно считать раскладное «крыло» столов-тумб или столов-книжек, выдвижное торцевое крыло.
Цельная центральная вставка раздвижного стола
Стол Спейс-6СТ фабрики Comfortstol
Составная вкладка — дополнительную площадь формирует не один (как в случае с цельной вкладкой), а несколько элементов. Частные случаи составных вкладок — торцевые выдвижные или те же «двойные крылья» столов-книжек.
Заметим, что в ряде случаев для формирования столешницы с помощью цельных и составных вкладок придется действовать исключительно с применением грубой физической силы. Такие элементы могут храниться как на пандусе под основной столешницей, так и отдельно. То есть «изымать» вкладку или вкладки из мест хранения и устанавливать на места использования, а затем повторять все операции в обратном порядке придется вручную. Но есть и более современные варианты, которые обеспечивают действительно комфортную трансформацию. Подробнее — в описании механизмов раздвижения.
Но есть и более современные варианты, которые обеспечивают действительно комфортную трансформацию. Подробнее — в описании механизмов раздвижения.
Составная торцевая вкладка из двух элементов
Стол Ramos от Avanti
Разрезная вкладка-бабочка — состоит из двух частей, соединенных петлями. В сложенном виде представляет собой «книжку». Зачастую (но не всегда) одно из преимуществ такого элемента состоит в менее трудоемкой фиксации по месту и обратно. Это возможно, когда используется поворотно-рычажный механизм, поэтому на весу (на руках) вставку держать не приходится. А самое сложное в процессе совсем не сложно — раскрыть или сложить «крылья».
Центральная вставка-бабочка
Стол B2303 бренда Gomlden
Все «раздвижные» вкладки могут располагаться практически в любом месте столешницы: по центру, у торца или торцов. Раскладные вставки-крылья — сбоку или по бокам. Место вставки зависит от примененных в изделии механизмов раздвижения.
Раздвижные механизмы трансформации
Оглавление раздела:
- Царговые столы.
- Механизм фронтслайд.
- Бесцарговые столы.
В царговых столах трансформация столешницы, как правило, выполняется только ее перемещением, в силу особенностей конструкции основание (подстолье) остается на месте.
В бесцарговых столах в трансформации участвуют опоры — при увеличении площади столешницы они сдвигаются относительно друг друга. Существуют промежуточные варианты трансформации типа «фронтслайд». Для сохранения прочности, надежности и устойчивости разложенного изделия могут применяться дополнительные опоры под столешницу.
Царговые столы
Несинхронные раздвижные механизмы с царговым креплением. Этот вариант в нынешнее время встречается редко, характерен для классических раздвижных столов с центральной вставкой. Трансформацию производят выдвижением полустолешниц поочередно — с одной, а затем с другой стороны.
Синхронные раздвижные механизмы трансформации царгового крепления более удобны — нужно потянуть лишь за одну часть столешницы, вторая выдвинется в противоположном направлении самостоятельно за счет тросового соединения.
Синхронный механизм можно опознать по наличию тросиков в механизме
Стол Бахус, Беларусь
Механизмы царгового крепления бывают внутренними и внешними. Внутренние совершенно не видны снаружи, их можно применять с материалом любой толщины. Но имеют жесткие ограничения по длине вставки. Механизмы внешнего крепления менее эстетичны, поэтому их чаще используют в моделях с декоративной фальш-царгой. Но зато такой механизм позволяет инсталлировать вставку чуть большего размера, чем для царговых столов. Как правило, в царговых столах используется одна цельная центральная вкладка шириной от 340 до 650 мм.
Фальш-царга закрывает механизм раздвижения
Стол-трансформер Версаль, пр-во Россия
Фронтслайд
Раздвижной механизм фронтслайд отличается от традиционного царгового и бесцаргового тем, что сдвигаются лишь две опоры с боковой царгой, а вкладка располагается не в середине, а у торца стола. Это решение позволяет увеличить длину столешницы до 640 мм.
Это решение позволяет увеличить длину столешницы до 640 мм.
Механизм «фронтслайд» — сдвигается не только столешница, но и опоры
Стол Lars от ItalGruppo
Бесцарговые столы
Многоколенные раздвижные механизмы бесцаргового крепления дают возможность изменять размер столешницы пошагово, укладывая нужное число вставок. Трансформацию выполняют с одной стороны, вытягивая опору (ножки) в горизонтальном направлении. Типичный пример применения — стол-консоль. Минус такого механизма — не всегда есть место для хранения вставок и необходимость использования дополнительной опоры. Плюс — возможность очень значительного увеличения длины столешницы — в несколько раз относительно стартового размера.
Многоколенный раздвижной механизм бесцаргового крепления
Стол-консоль Giant от Levmar, Россия
Чуть менее сложные синхронные раздвижные механизмы бесцаргового крепления используют не в консолях, а в больших обеденных столах. Они так же удобны, как синхронные царговые, но благодаря им можно использовать не одну, а несколько вставок, серьезно увеличивая площадь столешницы. С такими механизмами можно увеличить длину стола до 2000 мм. При этом составные вставки можно хранить под столешницей.
Они так же удобны, как синхронные царговые, но благодаря им можно использовать не одну, а несколько вставок, серьезно увеличивая площадь столешницы. С такими механизмами можно увеличить длину стола до 2000 мм. При этом составные вставки можно хранить под столешницей.
Синхронный раздвижной механизм бесцаргового крепления
Стол-трансформер Optimata 303S
Раздвижной несинхронный механизм торцевого подъёма столешницы с газлифтом. Трансформация выполняется вытягиванием и выталкиванием вверх вкладок с торцов стола. Не требует сколь-нибудь серьезных физических усилий, идеальный вариант для людей хрупкого телосложения. Одно из преимуществ — возможность увеличивать площадь как с двух, так и с одной стороны на 300 и 600 мм соответственно.
Вытяните дополнительную вкладку из-под столешницы и приподнимите — она займёт своё место
Широкий раздвижной стол Colombo от ItalGruppo
Раздвижной синхронный механизм с автоматическим подъемом вставки. Трансформацию выполняют вытягиванием полустолешницы с одной стороны до упора. Вторая половина изменяет положение синхронно, а центральная вставка самостоятельно поднимается вровень с основными. Все очень просто и под силу даже дамам субтильной конституции. Размер центральной вставки — порядка 300-320 мм.
Трансформацию выполняют вытягиванием полустолешницы с одной стороны до упора. Вторая половина изменяет положение синхронно, а центральная вставка самостоятельно поднимается вровень с основными. Все очень просто и под силу даже дамам субтильной конституции. Размер центральной вставки — порядка 300-320 мм.
Автоматический подъем вставки и синхронное раздвижение
Стол Anton от ItalGruppo
Раскладные механизмы трансформации
Классический раскладной механизм реализован в столах-книжках. В сложенном положении «крылья» столешницы висят по бокам стола. Для раскладывания стола их поднимают и устанавливают под них ножку-опору.
Сдвигово-раскладной механизм применяют со столешницами-книжками («двойная» разрезная столешница с соединением на петлях). В сложенном состоянии она лежит на раме. Трансформацию выполняют, сначала сдвигая сложенную столешницу в сторону, а затем «раскрывая» ее, как книгу.
Сдвигово-раскладной механизм трансформации столешницы
Стол ВМ50 фабрики Логарт
Поворотно-раскладной механизм тоже применяют для «сложенных» столешниц. В отличие от предыдущего варианта, здесь сложенную столешницу нужно сначала повернуть вокруг горизонтальной оси на опорной площадке, а затем разложить.
В отличие от предыдущего варианта, здесь сложенную столешницу нужно сначала повернуть вокруг горизонтальной оси на опорной площадке, а затем разложить.
Поворотно-раскладной механизм трансформации столешницы
Столик Агат 19.2, Россия
Раскладные механизмы, конечно, требуют чуть меньше сил, чем раздвижные с устанавливаемыми вручную вставками. Но больше, чем раздвижные с автоматической фиксацией вставок. Если столешница достаточно велика, при раскладывании и складывании вес будет чувствоваться. Также есть риск прищемить пальцы, с детства знакомый всем обладателям классического советского раскладного стола.
Минусы раскладных столов — недостаточно высокая эстетика как в сложенном, так и в разложенном виде. Видно петли. Кроме того, в разложенном виде столешницы недостаточно прочны и при возникновении избыточной нагрузки на края «идут на излом» — садиться и облокачиваться на край разложенной столешницы будет нельзя.
Механизмы подъема столешницы
Механизмы подъема столешницы можно разделить на механические, пневматические и электрические.
Электрические применяются редко, и в бытовых условиях не особо востребованы. В этой статье мы их не рассматриваем. Механические и пневматические по долговечности и надежности аналогичны, разница лишь в удобстве и простоте использования.
Рычажные подъемные механизмы могут быть чисто механическими с шарнирами, пружинами, а могут иметь в своем составе пневмоцилиндры (тогда они пневматические). Серьезных усилий не требуют, трансформация достаточно комфортна и безопасна.
Подъем столешницы за счет раскладывания опор реализован в столах Александрия или Агат-24.10 (внизу на фото). Систему типа Александрии мы не рекомендуем раскладывать женщинам и детям — нужна определенная физическая сила.
Подъем за счет раскладывания опор. Недостатки: люфты, расшатывание
Столик-трансформер Александрия
Подъем за счет подъема стального механизма опор
Столик-трансформер Агат 24.10
Механизм подъема ножничного типа реализован во всех изделиях с перекрещивающимися ножками (Х-образная форма опор). Подъем может осуществляться механическим и пневматическим способом.
Подъем может осуществляться механическим и пневматическим способом.
Механическая система, разумеется, требует больше физических сил, чем пневматическая, особенно при фиксации высоты с помощью винта (необходимо держа стол на весу, закрутить винт и зафиксировать высоту столешницы).
Для предупреждения повреждений напольных покрытий при подъеме и складывании столов с ножничными опорами в подавляющем большинстве случаев на ножках предусмотрены какие-либо защитные устройства. Это могут быть скрытые ролики, колесики, внешние ролики и пр. Один из весомых плюсов таких столов-трансформеров — очень малая высота в сложенном виде (у некоторых моделей всего порядка 30 см). Это дает возможность вообще убрать изделие с недостаточно высокой эстетикой с глаз долой после использования — спрятать стол в кладовке или шкафу.
Подъем ножничного типа (Х-форма опоры) плюс газлифт. Высота может составлять от 30 см
Стол В2293 бренда Gomlden
Подъемный механизм лифтового типа с газлифтом (пневмоцилиндром) не только удобен, но и эффектен. Например, стол с основанием-колонной, что вы видите на фото внизу, выглядит просто изумительно. Инженеры и дизайнеры предлагают чаще телескопические одноопорные и двухопорные варианты, причем опоры могут быть прямоугольного, круглого, овального сечения или даже выполнены в форме классических ножек. Газлифты, в зависимости от исполнения, могут обеспечивать шаговую или бесшаговую фиксацию столешницы по высоте. Такое решение не требует серьезных физических усилий.
Например, стол с основанием-колонной, что вы видите на фото внизу, выглядит просто изумительно. Инженеры и дизайнеры предлагают чаще телескопические одноопорные и двухопорные варианты, причем опоры могут быть прямоугольного, круглого, овального сечения или даже выполнены в форме классических ножек. Газлифты, в зависимости от исполнения, могут обеспечивать шаговую или бесшаговую фиксацию столешницы по высоте. Такое решение не требует серьезных физических усилий.
Подъемный механизм с газлифтом: удобно и легко
Стол-трансформер Optimata 312
Материалы для столов-трансформеров
Для изготовления столов-трансформеров используют абсолютно все, что только предлагает промышленность. Выбор обусловливается только вкусовыми и стилистическими предпочтениями и требованиями к экологичности. Заметим, что все материалы допущены для использования в быту, в жилых и хозяйственных помещениях. Но если экологичность занимает верхние строки в списке критериев выбора, скажем следующее.
Металл и стекло. Им можно отдать лидерство по экологичности. Придраться здесь просто не к чему. Материалы долговечны и поддаются освежению полировкой. Ну а ронять тяжелые и острые предметы нельзя на любую столешницу.
Дерево и шпон. Вопреки расхожему мнению, даже в 16-19 вв. именно массив дерева ценных или более-менее ценных пород использовался крайне редко. В подавляющем большинстве случаев речь шла об основании из дешевой древесины, фанерованной (шпонированной) более дорогими породами. Да, МДФ или ДСП не использовали, но, скорее, потому, что подобной технологии попросту не существовало. Сейчас древесные плиты на порядки безопаснее тех, что производились полвека назад, поэтому опасаться их не стоит. Шпонированные столы столь же, а в ряде случаев и более, красивы, чем изделия из массива.
Пластик. Синтетические материалы используют как в качестве мелких элементов фурнитуры, так и для создания прочнейшего и текстурированного покрытия плит МДФ или ДСП. Повторим: при соблюдении условий эксплуатации они безопасны.
подъёмные, раздвижные, откидные, поворотные, складные, выдвижные
В маленьких квартирах большой популярностью пользуется мебель-трансформер. Такие конструкции в собранном состоянии представляют собой один вид мебели, а в разобранном – другой. Удобный и практичный вариант – раскладной обеденный или письменный стол.
Разновидности механизмов трансформации столов
Стол, особенно обеденный, занимает много места. Он сам имеет довольно большую площадь, а расставленные вокруг стулья занимают еще больше пространства. Стационарная обеденная группа удобна при наличии отдельной столовой или большой кухни. Когда же зал объединяет кабинет, гостиную и столовую, нужны практичные решения.
Стол-трансформер – конструкция, которая может изменять площадь и положение столешницы. Различают их по количеству возможных трансформаций, механизмам раскладки и виду в собранном состоянии. Складной стол-книжка превращается в подобие узкого комода, которым, однако, пользоваться нельзя. Журнальная складная тумба в сложенном виде успешно играет роль журнального столика.
Положительно
57.14%
Предпочитаю стационарные
42.86%
Для маленькой дачи подойдет
0%
Проголосовало: 7
Модификация столешницы
Это наиболее старый и наиболее распространенный механизм. Благодаря простым манипуляциям поверхность столешницы можно увеличить в полтора раза. При этом высота стола не изменяется, ножки не переставляются. Такие столы называют раздвижными.
Существует немало модификаций.
Вставка – столешница разделена на 2 части. При необходимости можно раздвинуть обе части в стороны и вставить в середину дополнительное полотно. Вставка хранится отдельно в шкафу.
Поворотная центральная часть – механизм аналогичный, но вставка разделена на 2 части и находится под основной столешницей. Когда боковые части раздвигают, вставку вытаскивают наружу, разворачивают на 180 градусов и вставляют в проем.
Двойная столешница – сверху цельная поверхность, снизу под ней – разделенная на 2 части. При раскладке верхнюю часть снимают, нижнюю раздвигают в стороны и вставляют верхнее полотно в проем.
При раскладке верхнюю часть снимают, нижнюю раздвигают в стороны и вставляют верхнее полотно в проем.
Многоуровневый – принцип тот же: несколько столешниц находятся одна под другой. Они цельные, могут иметь разные размеры и выдвигаются с разных сторон. Получается не ровная, а многоуровневая конструкция. Чаще встречается у журнальных или кофейных столиков.
Раскладные
В этом случае в модели есть дополнительные опоры, которые позволяют увеличить площадь столешницы в 1,5 или 2 раза.
Стол-книжка – в сложенном состоянии представляет собой узкий глухой пенал, который использовать нельзя. При раскладе столешницы по обе или с одной стороны опорной конструкции поднимают и подпирают складными или выдвижными ножками и опорами.
Безцарговый стол – механизм раскладки представляет собой направляющие под столешницей. Перед тем как вставить центральную часть, раздвигают не 2 части поверхности, а 2 части стола вместе с ножками.
Откидной вариант – кухонный стол, в котором столешница в сложенном состоянии опущена к стене. При раскладке откидывается и подпирается опорной ножкой.
Площадь безцаргового стола можно увеличить в 3 раза, если под выдвижные боковые части подставить опорные ножки.
Вариант с подъемным механизмом
Стол с подъемным механизмом увеличивает площадь столешницы и изменяет ее высоту.
Складной – столешница и скрещенные опоры, которые заодно служат и подъемным устройством. Регулировка высоты не выполняется.
Подъемный журнальный – размеры столешницы не изменяются. Она целиком поднимается вверх. Так как величина поверхности остается небольшой, такую модель чаще используют стол для ноутбука, кофейный, десертный.
Раскладной подъемный – при раскладке столешница увеличивается в 2 раза и одновременно поднимается на нормальную высоту. Доводка до нужного уровня автоматическая. Получается полноценный обеденный стол, хотя и не совсем приглядный.
Получается полноценный обеденный стол, хотя и не совсем приглядный.
Дизайнерские модели трансформеров функциональны и красивы. Например, круглый стол, как бы состоящий из смещающихся относительно друг друга кругов.
Что учитывать при выборе стола-трансформера
Чтобы правильно подобрать изделие, нужно ответить самому себе на некоторые вопросы.
- Назначение – у обеденного стола столешница по площади должна соответствовать числу обедающих. Письменный стол для 1 человека куда меньше в размерах.
- Функциональность – полностью складной стол или стол-книжка в собранном виде не используется. Журнальная тумба в сложенном состоянии выполняет роль журнального столика, в разложенном – обеденного.
- Механизм обеденного стола-трансформера – они все просты в эксплуатации, но нужно учитывать нюансы. Если вставка от раздвижного стола хранится отдельно, ее нужно где-то сберегать и каждый раз вставлять и убирать своими руками.
- Грузоподъемность – механизм раскладки и опорная конструкция трансформера рассчитаны на определенную нагрузку. Грузоподъемность раздвижного стола, например, меньше чем раскладного, в котором под выдвинутые столешницы можно подставить опору.
- Дизайн – мебель должна соответствовать стилю интерьера.
 Письменный стол для 1 человека куда меньше в размерах.
Письменный стол для 1 человека куда меньше в размерах.Изготавливают трансформеры из самых разных материалов: дерева, хромированной стали, алюминия, стекла, пластика. Можно найти изделие на любой вкус.
Можно найти изделие на любой вкус.
Раздвижной
4.7
Откидной
4.5
Подъемный
4
Популярные вопросы
Чем проще механизм раскладки, тем дольше прослужит стол?
Не всегда верное утверждение. Сроки эксплуатации зависят не от простоты, а от устойчивости конструкции к нагрузкам. Механизм на газлифтах не просто облегчает подъем столешницы, а обеспечивает большое количество подъемов. Вариант на пружинах служит меньше, потому что пружина изнашивается быстрее.
Есть столы-трансформеры, в которых можно хранить вещи?
Выпускаются раскладные модели, предусматривающие ящики, и полки, и ниши для хранения. Пример такого решения – складная журнальная тумба. В ее основании обустраивают выдвижные ящики и полки.
Требует ли ухода механизм раскладки?
Да. Периодически следует осматривать направляющие, узел трансформации, очищать их от пыли и грязи. Хотя бы 1 раз в год нужно смазывать двигающиеся части машинным маслом.
Механизмы расширения стола
Воспользуйтесь фильтрацией для выбора механизмов расширения стола по расширению, высоте, длине или модели.

Товар успешно добавлен в корзину
В корзину
Товар успешно добавлен в ваш список желаний
Ваш список желаний
Уловка стола 95×30мм, оцинкованный, белый, 2 части
Код: 001312 Слева: > 5000 единиц
Цена продукта
0,09 €6 шт.
0,32 €
Сравнивать
| Покрытие: | без покрытия |
Механический фиксатор 60x20x1,2мм, оцинкованный, желтый
Код: 012230 Слева: < 500 единиц
Цена продукта
0,11 € / шт.
Сравнивать
| Высота, мм: | 1,2 |
Защелка стола Ø50мм, оцинкованная, белая, круглая, (2 детали)
Код: 001233 Слева: < 1000 набор
Цена продукта
€0,59/ установлен
Сравнивать
| Покрытие: | без покрытия |
Защелка стола 62х17мм, оцинкованная, белая, (2 детали)
Код: 000153 Слева: < 500 набор
Цена продукта
0,66 € / комплект
Сравнивать
| Покрытие: | без покрытия |
Защелка стола 65х35мм, оцинкованная, белый цинк
Код: 004950 Слева: < 500 набор
Цена продукта
0,74 € / комплект
Сравнивать
| Покрытие: | белый цинк |
Защелка стола 102x23x11мм, оцинкованная, белая, 2 части
Код: 004383 Слева: < 5000 набор
Цена продукта
0,85 € / комплект
Сравнивать
| Покрытие: | без покрытия |
Фиксатор механический 62×25,3×1,5мм, оцинкованный, желтый
Код: 005113 Слева: > 5000 единиц
Стоимость упаковки
€1,21
0,121 €/шт.
Сравнивать
| Высота, мм: | 1,5 |
| Покрытие: | белый цинк |
Штифт для подключения к столу Ø8×12мм, без покрытия, белый цинк,.
 ..
..Код: 000973 Слева: > 5000 набор
Стоимость упаковки
€1.331
0,1331 €/ комплект
Сравнивать
| Высота, мм: | 8 |
| Покрытие: | без покрытия |
Тормоз для расширения стола, оцинкованный, черный,.
 ..
..Код: 009284 Слева: 6 шт.
Стоимость упаковки
€1.331
0,6655 €/шт.
Сравнивать
| Высота, мм: | 12 |
| Покрытие: | без покрытия |
| Марка: | ДЕКСО |
Защелка стола 70х30мм, оцинкованная, белая, с фиксацией, (2.
 ..
..Код: 003110 Слева: < 5000 единиц
Цена продукта
1,53 € / шт.
Сравнивать
| Покрытие: | без покрытия |
Настольный складной трубодержатель 38x38x2,5, ø12мм, оцинкованный,.
 ..
..Код: 003866 Слева: 5 шт.
Цена продукта
1,6919 € / шт.
Сравнивать
| Высота, мм: | 42,5 |
| Покрытие: | белый цинк |
Держатель трубки для складного стола, серый, пластик, 4 отверстия, Ø16
Код: 003778 Слева: < 500 единиц
Цена продукта
1,7646 € / шт.
Сравнивать
| Высота, мм: | 16 |
| Покрытие: | без покрытия |
Защелка стола 138-150мм, без покрытия, нержавеющая сталь, (2 детали)
Код: 003824 Слева: < 500 единиц
Цена продукта
2,42 € / шт.
Сравнивать
| Покрытие: | без покрытия |
Комплект расширения стола
Код: 1000523 Слева: < 500 единиц
Стоимость упаковки
6,06 € / шт.
Сравнивать
Механизм расширения стола, Basic (P1) 650x455x35мм, без…
Код: 000451 Слева: < 500 набор
Цена продукта
€9,35 / комплект
Сравнивать
| Высота, мм: | 35 |
| Покрытие: | без покрытия |
| Удлинитель, мм: | 455 |
Обычная цена
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Добавить товар в корзину
Сообщите нам, какой продукт вам нужен
Если у вас есть особые требования к аналогичному продукту, заполните форму ниже и сообщите нам характеристики продукта, размеры или другие необходимые вам параметры. Принимая во внимание чертеж продукта внизу, сообщите нам более подробную информацию о требуемом продукте. Мы свяжемся с вами, чтобы обсудить возможности.
Принимая во внимание чертеж продукта внизу, сообщите нам более подробную информацию о требуемом продукте. Мы свяжемся с вами, чтобы обсудить возможности.
Используйте фильтрацию для выбора механизмов расширения стола по вылету, высоте, длине или модели
Наверх
Позиционирующие столы с шарико-винтовой передачей
Мехатроника
Высокоточные позиционирующие столы с приводом от шарико-винтовых пар
Механизмы позиционирования для точного и воспроизводимого движения машины
IKO разрабатывает и производит широкий ассортимент столов для позиционирования, приводимых в движение шарико-винтовыми парами, которые действуют как надежные механизмы позиционирования в различных областях применения, от оборудования для производства полупроводников до высокоточного оборудования.
ТУ ТЭ ТСЛ ТСЛХ ТХ ТК ТС/КТ ТМ
Столы для точного позиционирования (ТУ)
Наши прецизионные столы позиционирования TU представляют собой тонкие и компактные механизмы позиционирования с U-образной направляющей и направляющим столом. Стол находится внутри направляющей гусеницы и является частью полной линейной направляющей качения, состоящей из двух рядов стальных шариков большого диаметра, контактирующих с дорожками качения в четырех точках.
- Эта специальная конструкция обеспечивает высокую точность и высокую жесткость даже при изменяющихся и сложных нагрузках.
- Шесть типоразмеров доступны с шириной гусеницы от 25 до 130 миллиметров.
- Доступны дополнительные сильфоны для предотвращения проникновения пыли и обработка поверхности черным хромом для предотвращения коррозии.
- Длина стола, тип шарико-винтовой передачи, двигатель и датчики могут быть изменены в соответствии с потребностями конкретного применения.

Получить модель САПР
Конфигурация и предложение
Столы для прецизионного позиционирования (TE)
Наши столы для прецизионного позиционирования TE представляют собой легкие и компактные механизмы позиционирования, изготовленные из высокопрочного алюминиевого сплава и скользящего стола, который перемещается по U-образной направляющей.
- Масса стола на 40% меньше, чем у стандартной серии TU.
- Прецизионные шарико-винтовые пары обеспечивают надежное и точное позиционирование.
- Встроенные компоненты смазки C-Lube для длительного срока службы и работы без обслуживания до пяти лет или 20 000 километров пробега и более.
- ШВП, двигатели и датчики могут быть изготовлены по индивидуальному заказу в соответствии с потребностями конкретного применения.
- Предлагается множество размеров и стилей, включая модели с малой высотой сечения и столы с фланцами или без них.

Получить модель САПР
Конфигурация и предложение
Столы для точного позиционирования с длинным ходом (TSL)
Наши столы для точного позиционирования с длинным ходом представляют собой легкие механизмы для позиционирования с длинным ходом, которые имеют два ряда очень жестких линейных направляющих вместе с подвижным столом и станиной из высокопрочных алюминиевых сплавов. и прецизионные шариковые винты.
- Параллельная конструкция линейных направляющих обеспечивает высокую устойчивость, точность и грузоподъемность.
- Столы разных размеров и с различной длиной хода доступны при ширине стола от 90 до 220 миллиметров.
- Специализированные кронштейны XY доступны для 2-осевых приложений.
- Встроенные компоненты смазки C-Lube для длительного срока службы и работы без обслуживания до пяти лет или 20 000 километров пробега и более.
- Конструкция станины из алюминиевого сплава может быть настроена для достижения длины хода более 2000 миллиметров по запросу.

Получить САПР Модель
Настройка и расчет
Прецизионные столы для позиционирования с высокой жесткостью (TSLH)
Наши столы для прецизионного позиционирования с высокой жесткостью представляют собой чрезвычайно точные и надежные механизмы позиционирования, которые оснащены скользящими столами и станиной из чугуна для обеспечения высокой жесткости и превосходного гашения вибрации. Параллельная конструкция двух наборов линейных направляющих обеспечивает высокую стабильность и точность, а также высокую грузоподъемность.
- Четыре размера доступны с шириной стола от 120 до 420 миллиметров.
- Встроенные компоненты смазки C-Lube для длительного срока службы и работы без обслуживания до пяти лет или 20 000 километров пробега и более.
- Двухосевые столы возможны при объединении двух блоков LH.
- Доступны дополнительные сильфоны для предотвращения проникновения пыли и обработка поверхности черным хромом для предотвращения коррозии.
Получить модель САПР
Конфигурация и предложение
Сверхточные столы для позиционирования (TX)
Наши сверхточные столы для позиционирования обеспечивают сверхточную точность хода и высокую грузоподъемность благодаря использованию линейной роликовой направляющей Super X от IKO на жесткой чугунной станине. Усовершенствованная система управления с обратной связью, включающая линейный энкодер с разрешением 0,016 мкм, гарантирует превосходную точность позиционирования.
- Четыре размера доступны с шириной стола от 120 до 420 миллиметров.
- Встроенные компоненты смазки C-Lube для длительного срока службы и работы без обслуживания до пяти лет или 20 000 километров пробега и более.
- Опциональная смазка с низким пылеобразованием доступна для использования в чистых помещениях.
Получить модель САПР
Конфигурация и предложение
Прецизионные позиционирующие столы для чистых помещений (TC)
Наши прецизионные позиционирующие столы для чистых помещений разработаны для класса 3 ISO (Федеральный стандарт США 209). E Class 1), такие как производство полупроводниковых и жидкокристаллических дисплеев. Листы из нержавеющей стали и боковые крышки предотвращают попадание пыли, образующейся внутри стола, в чистую комнату.
E Class 1), такие как производство полупроводниковых и жидкокристаллических дисплеев. Листы из нержавеющей стали и боковые крышки предотвращают попадание пыли, образующейся внутри стола, в чистую комнату.
- Столы легкие, компактные и имеют минимальную высоту поперечного сечения всего 50 миллиметров.
- Основные компоненты изготовлены из высокопрочных, устойчивых к коррозии алюминиевых сплавов, нержавеющей стали и прецизионных шарико-винтовых пар.
- Ширина кровати варьируется от 50 до 86 миллиметров.
- Встроенные компоненты смазки C-Lube для длительного срока службы и работы без обслуживания до пяти лет или 20 000 километров пробега и более.
Получить модель САПР
Конфигурация и предложение
Компактные прецизионные столы для позиционирования (TS/CT)
Наши компактные прецизионные столы для позиционирования отличаются малой высотой сечения, высокой надежностью и высокой точностью, а также оснащены скрещенными роликовыми направляющими IKO в качестве направляющего механизма. Очень жесткий, гасящий вибрацию скользящий стол и станина из чугуна сочетаются с прецизионным приводным механизмом с шарико-винтовой передачей.
Очень жесткий, гасящий вибрацию скользящий стол и станина из чугуна сочетаются с прецизионным приводным механизмом с шарико-винтовой передачей.
- Доступны как одноосевые (TS), так и двухосевые (CT) модели для приложений с высокой точностью позиционирования, таких как прецизионные измерительные приборы и сборочные системы.
- Ширина стола составляет от 55 до 260 миллиметров для одноосных столов и от 55 до 350 миллиметров для двухосевых столов.
- Дополнительные роликовые направляющие с защитой от проскальзывания хорошо подходят для использования с вертикальной осью и высоким ускорением и замедлением.
Получить модель САПР
Конфигурация и предложение
Столы для позиционирования Micro Precision (TM)
Наши прецизионные микростолы для позиционирования представляют собой ультракомпактные механизмы, состоящие из микролинейной направляющей IKO L с шириной дорожки всего 2 мм и прецизионной шлифованной шарико-винтовой передачи диаметром 2 мм.
- Для всех основных компонентов используется коррозионностойкая нержавеющая сталь.
- Габаритные размеры составляют 20 миллиметров в высоту и 17 миллиметров в ширину.
- Максимальная скорость стола составляет 150 миллиметров в секунду при использовании шарико-винтовой передачи с высоким шагом и серводвигателя с высоким крутящим моментом.
- Скользящие столы доступны как в стандартной, так и в удлиненной версии с шаговыми двигателями или серводвигателями переменного тока.
- При необходимости доступны дополнительные датчики.
Получить модель САПР
Конфигурация и предложение
Письменный стол из красного дерева эпохи Регентства с механизмом скользящей высоты для продажи на 1stDibs
Товары похожие на Письменный стол из красного дерева периода Регентства с механизмом скользящей высоты
Письменный стол из красного дерева эпохи Регентства
Письменный стол из красного дерева Gillows периода Регентства 19-го века
Библиотечный стол раннего 19-го века английского Регентства с письменным слайдом
Письменный стол из красного дерева в стиле Регентства
Письменный стол из красного дерева позднего Регентства
2 843,25 долларов СШАО
Стол для письма или чтения из красного дерева позднего Регентства. На подвижной и регулируемой трещотке. На массивных точеных опорах. Включая скользящий механизм высоты. Высота может регулироваться до 51 дюйма.
На подвижной и регулируемой трещотке. На массивных точеных опорах. Включая скользящий механизм высоты. Высота может регулироваться до 51 дюйма.
Подробная информация
Размеры
Высота: 81,28 см (32 дюйма) Ширина: 88,9 см (35 дюймов) Глубина: 53,34 см (21 дюйм)
Стиль
(из периода)
(из период
.
Place of Origin
Англия
ПЕРИОД
РАБОТАЯ 19-го века
Дата производства
1825-1830
- .0005
Справочный номер
Продавец: 79421STDIBS: LU1032888821933
View INVOICE
Доставка и возврат
1stdibs. сделать это правильно. Узнать больше
Части этой страницы были переведены автоматически. 1stDibs не может гарантировать точность переводов. Английский язык является языком по умолчанию на этом веб-сайте.
О продавце
Расположен в Питерборо, Великобритания
Золотой продавец
Эти проверенные экспертами продавцы имеют высокие оценки и неизменно превосходят ожидания клиентов.
Основано в 1957 году
1stdibs Seller с 2014 года
649 Продажи на 1stDibs
Типичное время отклика: 1 час
Подробнее от этого продавца
ПИСЬМЕННЫЙ ПЕРИОНТ И РЕЧЕТА
Расположенный в Питере, Северный,
Период.
.0005
Хороший стол для письма/чтения эпохи Регентства с подъемным центром прямоугольного сечения, арочной платформой очень хорошей формы, заканчивающейся оригинальными туфлями и колесиками, столешницей необычно…
Материалы
Красное дерево
Стол для письма и чтения периода Регентства
4 435 долларов США
Стол для чтения и письма периода Регентства
Расположен в Питерборо, Нортгемптоншир
Складной карточный столик эпохи Регентства, который также открывается на стол для чтения и письма необычно малых размеров.
Тонко вырезанная и точеная ножка над основанием платформы на резной…Категория
Антикварные столы и письменные столы эпохи британского Регентства XIX века
Стол для чтения и письма эпохи Регентства
3 298 долларов США
Письменное дерево эпохи Георга III из красного дерева Таблица
Расположен в Питерборо, Нортгемптоншир
Исключительно красивый письменный стол из красного дерева эпохи Георга III с кожаной вставкой для письма с закругленными краями кверху, тонкими сужающимися квадратными опорами с коробчатыми секциями, инкруст…
Категор …
Материалы
Красное дерево
Письменный стол из красного дерева периода Георга III
29 569 долларов США
Библиотечный стол из красного дерева с шестью ящиками позднего периода Регентства
Расположен в Питерборо, Нортгемптоншир.
Библиотечный стол из красного дерева с шестью ящиками периода позднего Регентства.
На точеных опорах, заканчивающихся оригинальными башмаками и колесиками. Хороший общий цвет. Столешница изношена, но в отличном состоянии…Категория
Старинные столы и письменные столы эпохи английского Регентства начала XIX века
Библиотечный стол из красного дерева с шестью ящиками позднего Регентства
14 102 долл. США
ПериодЧиппендейл Письменный стол
Расположен в Питерборо, Нортгемптоншир.
Элегантный стол для чтения и письма из красного дерева периода Чиппендейла на очень хорошо вырезанной центральной сужающейся опоре, блок над ножками со вставными «перфорированными» панелями, двойная храповая регулировк…
Категория
Антиквариат Британские письменные столы и письменные столы Чиппендейла 18-го века и ранее
Материалы
Красное дерево
Период Чиппендейла Стол для чтения или письма из красного дерева
2 956 долл.
СШАРабочий / письменный стол из красного дерева периода Георга III
Расположен в Питерборо, Нортгемптоншир
Самый привлекательный рабочий / письменный стол из красного дерева периода Георга III. Почти наверняка Гиллоуз из Ланкастера. Центральная секция с подъемной крышкой, открывающей храповую поверхность для письма …
Категория
Старинные английские письменные столы и письменные столы эпохи Георга III 1780-х годов
Материалы
Красное дерево
Период Георга III Красное дерево Рабочий / письменный стол
4 435 долларов США
 Тонко вырезанная и точеная ножка над основанием платформы на резной…
Тонко вырезанная и точеная ножка над основанием платформы на резной… На точеных опорах, заканчивающихся оригинальными башмаками и колесиками. Хороший общий цвет. Столешница изношена, но в отличном состоянии…
На точеных опорах, заканчивающихся оригинальными башмаками и колесиками. Хороший общий цвет. Столешница изношена, но в отличном состоянии… США
СШАВам также могут понравиться
Письменный стол из красного дерева Regency
Расположен в Эссексе, Массачусетс ножки и ролики.
Категория
Антикварные столы и письменные столы эпохи Регентства в английском стиле начала XIX века
Материалы
Красное дерево
Письменный стол из красного дерева Регентства
7 500 долларов США
Письменный стол из красного дерева Gillows периода Регентства XIX века
Находится в Лондоне, Великобритания
Прекрасный письменный стол начала XIX века из красного дерева с шестью ящиками, кожаным верхом с тростниковой окантовкой и двумя подставками для чтения. над вышитыми бисером ящиками по бокам …
над вышитыми бисером ящиками по бокам …
Категория
Антикварные столы и письменные столы эпохи Британского Регентства начала XIX века
Материалы
Красное дерево, кожа
Письменный стол Gillows из красного дерева периода Регентства XIX века
30 707 долларов США
Библиотечный стол эпохи Регентства Англии начала XIX века с письменным слайдом
Расположен в Чарльстоне, Южная Каролина
Этот большой библиотечный стол был изготовлен в Англии в первой половине 19-го века. Этот стол из красного дерева с коричневой кожаной столешницей и восемью ящиками (четыре искусственных). Один …
Категория
Антикварные столы английского периода Регентства начала XIX века
Материалы
Кожа, красное дерево
Библиотечный стол в стиле английского Регентства начала XIX века с письменным столиком
14 500 долл. США
США
Письменный стол из красного дерева в стиле Регентства
поднимается на вывернутые ноги. Задняя часть закончена, но нет ящиков.Категория
Антикварные столы и письменные столы эпохи английского Регентства начала XIX века
Материалы
Кожа, красное дерево
Письменный стол из красного дерева в стиле Регентства
2 950 долларов США
Письменный стол из красного дерева в стиле позднего Регентства
Расположен в Нью-Йорке, штат Нью-Йорк
Письменный стол из красного дерева в стиле позднего Регентства, ок. 1835 г., с прямоугольной столешницей со вставкой из кожи бутылочно-зеленого цвета, над двумя короткими ящиками с патентованными замками, на четырех точеных сужающихся тростниковых ножках на…
Категория
Старинные столы и письменные столы эпохи Регентства середины XIX века
Материалы
Красное дерево
Письменный стол из красного дерева эпохи Регентства
2 800 долларов США
Небольшой письменный стол из красного дерева эпохи Регентства
Расположен в Бедфордшире, Великобритания
Высококачественный письменный стол из красного дерева периода Регентства
Имея два встроенных ящика для фриза с оригинальным
Точеные ручки приподняты на элегантном конце с х-образной рамкой
Опоры в оригинальном латунном корпусе. ..
..
Категория
Антикварные приставные столики эпохи английского Регентства начала XIX века
Материалы
Красное дерево
Маленький письменный стол из красного дерева эпохи Регентства
6 653 долл. США
Метаморфический письменный/книжный стол периода Регентства
Расположенный в Гудзоне, штат Нью-Йорк
Этот прекрасный классический стол эпохи Регентства демонстрирует все признаки лондонского фирменного кабинета. Пропорции отличные, не слишком большие, но они имеют вес и выглядят мужественными и …
Категория
Антикварные столы и письменные столы эпохи Регентства XIX века
Материалы
Латунь
Метаморфический письменный/книжный стол эпохи Регентства
5 150 долларов США
Письменный стол из красного дерева эпохи Регентства
Находится в Эссексе, Массачусетс , приподнятые на круглых конических ножках и роликах.
Категория
Антикварные столы и письменные столы начала XIX века
Материалы
Красное дерево
Письменный стол из красного дерева Regency
6 800 долларов США
Письменный стол из красного дерева Regency с инкрустацией
Расположен в Эссексе, Массачусетс. С прямоугольной столешницей с латунной галереей 3/4 и поверхностью для письма из черной кожи. Fr…
Категория
Антикварные столы и письменные столы эпохи английского Регентства начала XIX века
Материалы
Красное дерево
Письменный стол Regency из красного дерева и инкрустацией
8 950 долларов США
Письменный стол Regency из красного дерева и черного дерева
Находится в Эссексе, Массачусетс. концы в форме лиры, ножки сабли и медные ролики.
Категория
Старинные письменные столы и письменные столы эпохи Регентства 1810-х гг.

Материалы
Латунь
Письменный стол из красного дерева и черного дерева Регентства
US 12 000 долл. США
Недавно просмотр
View MoreОбещание 1stDibs
Узнайте больше
. Доставка
Дополнительные способы просмотра
Письменный стол Regency Антикварная мебель
Письменный стол Regency
Письменный стол Regency 2
Рейсинс письменная таблица стола
Degency Desk England
Стол письма с регентством
Период регенсии Таблица
Столы регенции Mahogany
Античный слайд
Регенпия Период.
Письменный стол из красного дерева Regency 19th Century
Передвижной стол
Высокие столы
Просмотреть все Популярные письменные и письменные столы Поиски
Механизмы для раздвижных столов — Roble.
 Store
StoreЭкологическая мебель «Roble · Store» гарантирует высокое качество и долговечность в аутентичном скандинавском стиле. Все виды мебели изготовлены из массива дерева марок NordicStory и loftstory , которые имеют долговечные привычки комфорта и дают много качества жизни. Раздвижные обеденные столы в нашей коллекции выполнены из элегантного массива дуба и отвечают требованиям, прежде всего, надежной безопасности в вашем доме, благодаря благородным материалам и скандинавскому дизайну.
Так как мы специализируемся на всех видах дубовой мебели, у нас есть несколько механизмов складных столов. Мы поможем вам подобрать стол с максимально надежным и удобным механизмом, который максимально соответствует вашим требованиям дома.
Перед тем, как выбрать один Раздвижной обеденный стол , очень важно оценить и проанализировать доступное пространство в соответствии с требуемыми размерами.
При покупке детально измерьте площадь, чтобы рассчитать размер необходимого вам стола. Необходимо учитывать увеличенную длину стола. Советуем оставить не менее 90 см дополнительного пространства с каждой стороны стола для размещения комплекта обеденных стульев.
Необходимо учитывать увеличенную длину стола. Советуем оставить не менее 90 см дополнительного пространства с каждой стороны стола для размещения комплекта обеденных стульев.
Типы раздвижных обеденных столов
Во-первых, давайте взглянем на удлинители на самой раме стола. Вы можете выбрать в соответствии с различными формами разнообразного дизайна раздвижных обеденных столов, но мы также предлагаем некоторые варианты удлинителей, которые сегодня больше всего представлены на рынке.
Механизм выдвижения «Бабочка»
Сам механизм выдвижения «Марипоса» очень прост и удобен в использовании и надо отметить, что он является лидером продаж, среди всех самых современных столов этого типа, которые доступно сегодня. Он одобрен и надежен. Ножки стола зафиксированы. Чтобы разложить стол, двигаем доску и достаем нашу «Бабочку». При таком механизме площадь поверхности увеличивается на 30-40%. С обеих сторон стола под столешницей находятся английские булавки. Это два пластиковых рычага, которые в целях безопасности не дадут столу открыться. Стол собран с заводским механизмом «Баттерфляй». Заказчик починит ноги, не более того.
С обеих сторон стола под столешницей находятся английские булавки. Это два пластиковых рычага, которые в целях безопасности не дадут столу открыться. Стол собран с заводским механизмом «Баттерфляй». Заказчик починит ноги, не более того.
Как вы можете увидеть Раздвижные столы конструкции: Moby, Scandi 3, Scandi 2, Франция Они устанавливаются с помощью механизма «бабочка» того же типа.
Extendeble Dining Table «France»
Столовый стол «Escandi 3»
Стол для обеда с механизмом расширения «Баттерфля и идеальный способ принять столько гостей, сколько вы хотите, не занимая много места, а когда вы открываете его, вокруг него может сидеть больше людей. Преимуществом данного поворотного механизма является высокая скорость разборки стола. Современное приспособление и технология изготовления позволяют покупателям легко раскладывать стол, затрачивая на это совсем немного своего времени, не более минуты.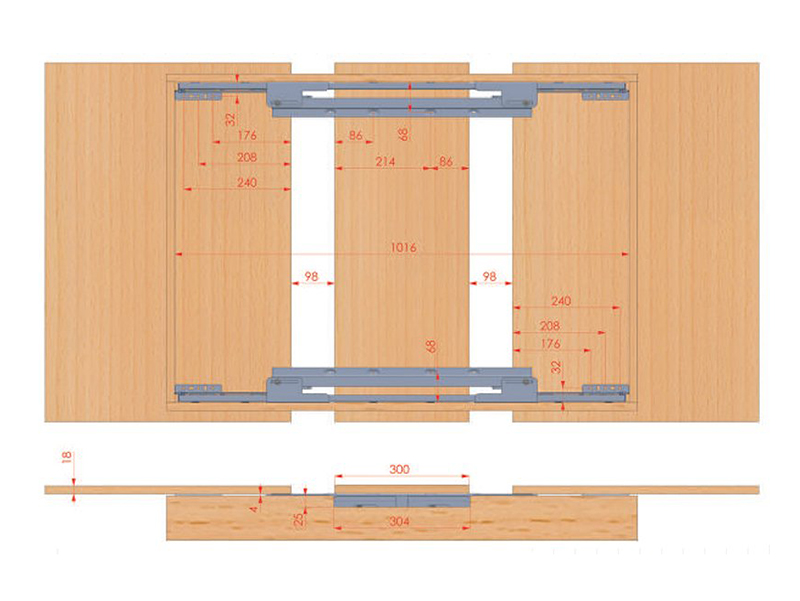 Продуманный механизм расширения складывается и поворачивается, чтобы поместиться внутри стола, скрывая себя, когда он не используется. Внутренние стержни, удерживающие удлинитель, просты в использовании, как и Раздвижной стол «Moby» благодаря этому он более прочный. С механизмом расширения «Бабочка» вам никогда не придется беспокоиться о хранении дополнительных листов, так как они легко складываются поверх стола.
Продуманный механизм расширения складывается и поворачивается, чтобы поместиться внутри стола, скрывая себя, когда он не используется. Внутренние стержни, удерживающие удлинитель, просты в использовании, как и Раздвижной стол «Moby» благодаря этому он более прочный. С механизмом расширения «Бабочка» вам никогда не придется беспокоиться о хранении дополнительных листов, так как они легко складываются поверх стола.
Раздвижной обеденный стол «Moby»
Стол с центральным удлинением нижняя часть внутри стола. Это классический механизм, проверенный многолетней практикой. Ножки стола остаются неподвижными. Чтобы расширить поверхность стола, столешницу перемещают и помещают центральную часть. С обеих сторон стола под столешницей находятся английские булавки. Это два пластиковых рычага, которые в целях безопасности не дадут столу открыться.
Раздвижной обеденный стол «Гарольд»
Будучи практичным и гибким в использовании, центральное выдвижное лезвие позволяет удвоить количество посадочных мест. Дополнительный лист держится под рукой, под столом, когда он не используется. Поэтому, если вам нужно дополнительное место для приема пищи, поместите центральную надставку, которую вы найдете внутри складного стола.
Дополнительный лист держится под рукой, под столом, когда он не используется. Поэтому, если вам нужно дополнительное место для приема пищи, поместите центральную надставку, которую вы найдете внутри складного стола.
Выдвижной механизм
Стол «Марси» уникален. Чтобы разложить его, мы тянем за ножки, которые имеют колесики и легко перемещаются. Внутри стола находится выдвижная часть, которая фиксируется на столешнице. Изготовлен из массива дерева, включая механизм, который надежен и не ломается. Он идеально подходит для небольших помещений, его можно поставить рядом со стеной, потому что он не круглый.
Раздвижной обеденный стол «Marsi»
Продуманный дизайн означает, что стол не имеет шарниров, когда вы используете его без расширения. Ножки стола всегда находятся в углах стола, даже когда стол выдвинут, поэтому под столом достаточно места для стульев. Один человек может быстро и аккуратно накрыть стол перед приходом гостей на большую вечеринку. Дополнительный лист держится под рукой, под столом, когда он не используется.
Дополнительный лист держится под рукой, под столом, когда он не используется.
Поворотно-складной механизм
En Oak.Store У нас есть только один стол с поворотным механизмом — «Mini 2». Поэтому при раскладывании меняется ширина стола, а в остальных случаях удлиняется только длина стола. Чтобы разложить стол, вы должны повернуть доску и открыть ее, как если бы это была книга. Это своеобразный стол, поэтому столешница не фиксируется застежками. Будучи простой таблицей, это очень модно. Поверните стол на 180 градусов, чтобы развернуть другую его часть. Это технически очень удобный и простой в использовании вариант, но его конструкция не отличается изысканностью. Выберилучшее для себя!
Стол обеденный раздвижной «Мини 2»
Столешница двуспальная
Уникальность такого предмета в простоте механической части. В зависимости от складывания толщина стола увеличивается почти вдвое. Чтобы его разложить, достаточно поднять оба крыла, потянув в стороны. Центральная часть доски помещается в свободное пространство и готова к использованию. Поверхность такого стола увеличивается почти на 50%. Вы можете использовать только один из них или оба. Наши Стол «Мини 1» изготовлен из массива дерева, он небьющийся, длинный и узкий. В узких кухнях идеально размещать его рядом со стеной.
Чтобы его разложить, достаточно поднять оба крыла, потянув в стороны. Центральная часть доски помещается в свободное пространство и готова к использованию. Поверхность такого стола увеличивается почти на 50%. Вы можете использовать только один из них или оба. Наши Стол «Мини 1» изготовлен из массива дерева, он небьющийся, длинный и узкий. В узких кухнях идеально размещать его рядом со стеной.
Extendemable Dining Table «Mini 1»
Другие самые продаваемые таблицы из Roble Store
Столовая Table L2
IS Ase Aceply Asception Asception Adensense Adensense Adension. Немецкий бренд Poetiker . Чтобы разложить стол, аккуратно отодвигаем его борт в сторону и снимаем выдвижную удлиняющую часть. С обеих сторон стола под столешницей находятся английские булавки. Это два пластиковых рычага, которые в целях безопасности не дадут столу открыться. Элегантная особенность стола в том, что в сложенном состоянии посередине нет шарниров. На первый взгляд не кажется, что это раздвижной стол, однако он раскладывается за 2 минуты.
На первый взгляд не кажется, что это раздвижной стол, однако он раскладывается за 2 минуты.
Раздвижной обеденный стол «L2»
Стол XL
Это наш флагманский продукт. Столешница и ножки двигаются по этому столу. Это стол с тремя раздвижными листами. Все хранится внутри. Он не требует дополнительного места для хранения. Его можно использовать в закрытом виде и/или с 1, 2 или 3 развернутыми листами. В разложенном положении стол имеет 4 ножки; При раскладывании створок из внутренней части основных ножек автоматически выходят 2 дополнительные ножки. Максимальная длина 2 метра 80 см. Мы называем это большим семейным столом. Механизм немецкого бренда Poetiker плавно двигается благодаря колесикам на ножках.
Раздвижной обеденный стол «XL»
Mauritz 1 обеденный стол
Мне было интересно, понравится ли нашим клиентам такой стол, но мы оказались правы. Это исключительный стол. Стиль деревенский: ножки мощные и крепкие. Механизм в старинном стиле, возможно, не такой удобный, как современные, также имеет деревянную фурнитуру, которая хранится отдельно и НЕ ХРАНИТСЯ ВНУТРИ СТОЛА. Это единственная модель, которая не позволяет хранить дополнительный лист под столом. Мы знаем, что это не совсем удобно, но в то же время стол в закрытом положении выглядит красиво и показывает, что он изысканного качества и очень прочный. Массивные ножки и деревенский дизайн в моде в Испании. Он включает в себя удлинитель, но конструкция позволяет устанавливать его с любой стороны. Зазоры закрываются заглушками, которые открываются, когда необходимо поставить удлинитель.
Стиль деревенский: ножки мощные и крепкие. Механизм в старинном стиле, возможно, не такой удобный, как современные, также имеет деревянную фурнитуру, которая хранится отдельно и НЕ ХРАНИТСЯ ВНУТРИ СТОЛА. Это единственная модель, которая не позволяет хранить дополнительный лист под столом. Мы знаем, что это не совсем удобно, но в то же время стол в закрытом положении выглядит красиво и показывает, что он изысканного качества и очень прочный. Массивные ножки и деревенский дизайн в моде в Испании. Он включает в себя удлинитель, но конструкция позволяет устанавливать его с любой стороны. Зазоры закрываются заглушками, которые открываются, когда необходимо поставить удлинитель.
Раздвижной обеденный стол «Mauritz 1»
Советы по выбору раздвижного стола
Перед покупкой необходимо учесть следующее:
- Измерить пространство. Важно проанализировать свободное место для складного стола и посмотреть, как он впишется в дизайн интерьера. Если правильно подобрать габариты, изделие всегда можно использовать в собранном виде и удобно сложить стол к приходу гостей. И не стоит платить высокую цену за огромную доску, которая не понравится.
 Если правильно подобрать габариты, изделие всегда можно использовать в собранном виде и удобно сложить стол к приходу гостей. И не стоит платить высокую цену за огромную доску, которая не понравится.
Если правильно подобрать габариты, изделие всегда можно использовать в собранном виде и удобно сложить стол к приходу гостей. И не стоит платить высокую цену за огромную доску, которая не понравится.- Оценить функциональность продукта. Необходимо проанализировать, достаточно ли увеличения поверхности доски во время посиделок с семьей и друзьями. Вы также можете увидеть некоторые дополнительные возможности, удобные в использовании, такие как ящики для хранения столовых приборов, небольшие полочки для салфеток и других мелочей.
- Проверьте качество материалов и аксессуаров, из которых изготовлен стол. Если компоненты недостаточно надежны, конструкция может быстро разрушиться. Так что не нужно гнаться за дешевизной и тратить деньги на товары с коротким сроком службы.
Чем больше вы думаете о покупке раздвижного обеденного стола, тем дольше он прослужит, радуя хозяев и гостей возможностью приятно провести время за обедом и застольными беседами. По этой же причине интернет-магазин Oak.Store делает все возможное, чтобы покупатель мог купить за свои деньги самое лучшее. Не стесняйтесь обращаться к нашему торговому представителю, вместе с профессиональным менеджером по продажам они могут подробно проконсультировать вас по каждой модели изделия, мы также в вашем распоряжении, чтобы объяснить преимущества и недостатки, дать совет, как лучше всего установить мебель в интерьере. Сотрудничество с нами сделает Ваш дом уютным и комфортным для повседневной жизни!
По этой же причине интернет-магазин Oak.Store делает все возможное, чтобы покупатель мог купить за свои деньги самое лучшее. Не стесняйтесь обращаться к нашему торговому представителю, вместе с профессиональным менеджером по продажам они могут подробно проконсультировать вас по каждой модели изделия, мы также в вашем распоряжении, чтобы объяснить преимущества и недостатки, дать совет, как лучше всего установить мебель в интерьере. Сотрудничество с нами сделает Ваш дом уютным и комфортным для повседневной жизни!
Посетите наш выставочный зал по адресу: Барселона, Avenida Diagonal 352, 08013
7 советов по выбору раздвижных столов: форма, материал, механизм раскладки
Карина_Вайло | 29.10.2018 | Мебель для гостиной, Мебель для кухни | 6 722 просмотра | комментариев нет
Содержание статьи
Стол является неотъемлемым элементом кухни, а иногда и гостиной. Именно без этого предмета интерьера трудно будет представить любое застолье. Так, его размеров должно хватить для комфортного размещения всех членов семьи и гостей. Учитывая то, что площадь помещений в обычных квартирах не отличается особой вместительностью, подогнать стол достаточной длины и ширины может быть проблематично. Разумной альтернативой и настоящим спасением в таких случаях станет приобретение столов-трансформеров. Это современное решение позволит не только существенно сэкономить драгоценное пространство, но и подобрать предмет мебели, идеально вписывающийся в интерьер. В этой статье мы дадим 7 советов по выбору раздвижных столов , рассмотрим самые популярные формы столешницы, материал изготовления и типовые механизмы компоновки. Мы расскажем, на что нужно обратить внимание при выборе и как правильно эксплуатировать складные конструкции, чтобы продлить срок их службы.
Так, его размеров должно хватить для комфортного размещения всех членов семьи и гостей. Учитывая то, что площадь помещений в обычных квартирах не отличается особой вместительностью, подогнать стол достаточной длины и ширины может быть проблематично. Разумной альтернативой и настоящим спасением в таких случаях станет приобретение столов-трансформеров. Это современное решение позволит не только существенно сэкономить драгоценное пространство, но и подобрать предмет мебели, идеально вписывающийся в интерьер. В этой статье мы дадим 7 советов по выбору раздвижных столов , рассмотрим самые популярные формы столешницы, материал изготовления и типовые механизмы компоновки. Мы расскажем, на что нужно обратить внимание при выборе и как правильно эксплуатировать складные конструкции, чтобы продлить срок их службы.
Перед покупкой
Прежде чем отправиться на поиски своего идеального стола, вы должны понимать, что действительно качественная мебель может прослужить не один год. Если вы не из тех, кто привык каждые пару лет менять интерьер, а заодно и большую часть мебели, рекомендуем внимательно отнестись к выбору.
Если вы не из тех, кто привык каждые пару лет менять интерьер, а заодно и большую часть мебели, рекомендуем внимательно отнестись к выбору.
- В первую очередь необходимо определиться с необходимыми размерами. Что значит размеры стола в разложенном виде. Для более рационального использования столешницы гостей рекомендуется расставлять по всему периметру стола, а значит, его необходимо отодвинуть от стены к центру комнаты. Для свободного перемещения со всех сторон должно оставаться минимум 70-80 см;
- Обратите внимание, что при выборе размера стол также влияет на площадь комнаты. Раздвижной стол может быть не только кухонным, но и журнальным. Последний в сложенном состоянии имеет компактные размеры, а при необходимости его центральная часть поднимается вверх с помощью специальных механизмов и раскладывается в стороны. Таким образом, он увеличивается не только в длину, но и в высоту, и превращается в полноценный стол. Поэтому вспомните, сколько гостей вы принимаете в среднем за праздники и можно ли оставаться в гостиной. Особенно это актуально для небольшой семьи, которой вполне достаточно небольшого обеденного стола;
- Для комфортного использования требуется не менее 60 см столешницы на человека и 45 см в ширину. На фото ниже оптимальная схема рассадки гостей и необходимый размер в зависимости от количества человек;
- Учитывая средний рост взрослого человека, столы имеют стандартную высоту 75 см. Однако, учитывая индивидуальные потребности, возможно изготовление столов меньшей или большей высоты;
- Обратите внимание на соответствие внешнего вида изделий общему стилю помещения. Например, если интерьер выполнен в одном из современных стилей, где отдается предпочтение минимализму и использованию современных материалов, то деревянный стол в классическом стиле будет резко выделяться и казаться неуместным;
- Также сразу уточняйте в магазине о возможности подбора комплекта стульев к понравившейся модели. Многие производители сразу предлагают готовые обеденные группы. Это самый выгодный вариант как с точки зрения дизайна, так и в финансовом плане. После покупки нестандартного изделия у вас могут возникнуть трудности с выбором стульев или мягкого уголка.
 Особенно это актуально для небольшой семьи, которой вполне достаточно небольшого обеденного стола;
Особенно это актуально для небольшой семьи, которой вполне достаточно небольшого обеденного стола; Это самый выгодный вариант как с точки зрения дизайна, так и в финансовом плане. После покупки нестандартного изделия у вас могут возникнуть трудности с выбором стульев или мягкого уголка.
Это самый выгодный вариант как с точки зрения дизайна, так и в финансовом плане. После покупки нестандартного изделия у вас могут возникнуть трудности с выбором стульев или мягкого уголка.10 преимуществ и 1 недостаток столов-трансформеров
Раздвижные столы действительно можно назвать лучшим вариантом для использования в стесненных условиях. А в современных интерьерах, где особое внимание уделяется функциональности и эргономике, чаще всего используется мебель-трансформер. Кроме того, в процессе эксплуатации вы обязательно найдете еще несколько Преимущества Похожие конструкции:
- Эстетика. Дизайн раздвижных столов выполнен на высшем уровне и позволяет приобрести не просто предмет мебели, а настоящее украшение любого интерьера;
- Понятный механизм компоновки, который вы изучите на интуитивном уровне. Для того, чтобы быстро разобраться, как обращаться со столом, не нужно долго и мучительно изучать инструкцию;
- Простота использования. Даже самые сложные механизмы раскладки рассчитаны на минимальные физические усилия. Поэтому с задачей справится и хрупкая девушка, и подросток;
- Значительное увеличение размеров. Некоторые модели раздвижных столов способны увеличиваться в длину втрое, а то и вчетверо. Крупные модели в разложенном виде могут достигать длины около 4 метров;
- Регулировка нескольких параметров одновременно. Многие современные модели имеют возможность изменять не только длину столешницы, но и ее высоту. Это позволяет максимизировать настройки стола в соответствии с вашими потребностями;
- Повышенный комфорт. Конфигурация современных моделей продумана таким образом, что какой бы способ раскладки вы ни выбрали, вам всегда будет достаточно места для ног. Такие важные мелочи несомненно значительно повышают удобство использования;
- Различные материалы. Можно найти столы как из традиционных материалов, таких как ДСП, МДФ или дерево, так и совершенно нетипичные. Например, дорогие конструкции дополняют изящные столешницы из камня или керамической плитки. Это дает возможность не только подобрать модель в полном соответствии с кухонным гарнитуром, фартуком или другими предметами мебели, но и выбрать из другого ценового сегмента. Таким образом, даже при самом скромном бюджете найдется оптимальное предложение;
- Разнообразие форм и цветов. Это также позволяет упростить выбор;
- Высокая надежность и стабильность. Несмотря на то, что конструкция состоит из нескольких элементов и направляющего механизма, в разложенном виде по прочности она не уступает обычным цельным моделям. Раздвижные столы также могут выдерживать значительные нагрузки;
- При правильном и бережном обращении срок службы столов-трансформеров может достигать 10-15 лет. Согласитесь, это весьма внушительные цифры, учитывая особенности конструкции.
 Даже самые сложные механизмы раскладки рассчитаны на минимальные физические усилия. Поэтому с задачей справится и хрупкая девушка, и подросток;
Даже самые сложные механизмы раскладки рассчитаны на минимальные физические усилия. Поэтому с задачей справится и хрупкая девушка, и подросток; Это дает возможность не только подобрать модель в полном соответствии с кухонным гарнитуром, фартуком или другими предметами мебели, но и выбрать из другого ценового сегмента. Таким образом, даже при самом скромном бюджете найдется оптимальное предложение;
Это дает возможность не только подобрать модель в полном соответствии с кухонным гарнитуром, фартуком или другими предметами мебели, но и выбрать из другого ценового сегмента. Таким образом, даже при самом скромном бюджете найдется оптимальное предложение;ТО К недостаткам можно отнести только высокую стоимость. Однако при стольких преимуществах и столь длительном сроке службы можно сказать, что он полностью оправдан.
Виды выдвижных механизмов
Столы-трансформеры могут иметь различные типы механизмов раскладки. Наиболее долговечными и наиболее распространенными являются следующие варианты.
Наиболее долговечными и наиболее распространенными являются следующие варианты.
Стол раздвижной с выдвижным ящиком и выдвижным центром
Царьга — это массивное основание или каркас, к которому крепится столешница. Одной из самых популярных и простых конструкций столов с ручной раскладкой является модель с выдвижной центральной частью. В собранном виде по центру столешницы имеется стыковочный шов, а под самой столешницей отсек для размещения скользящая часть. Для того чтобы разложить такой стол, необходимо раздвинуть обе половины столешницы в стороны, а затем снять центральную часть и вставить ее в пазы. Затем отодвиньте боковые половинки до характерного щелчка. Это означает, что все части конструкции фиксируются в правильном и стационарном положении. При таком способе раскладки длина стола может увеличиваться на 1/3 или 1/2 длины в сложенном виде.
Аналогичные модели имеют следующие преимущества:
- Стоит отметить, что это одна из самых прочных, долговечных и надежных конструкций;
- Большинство столов сделаны так не просто так, а потому что это решение действительно проверено временем. Обратите внимание, что старые советские столы, которые умели раскладывать, имели именно такой каркас.
- Как в собранном, так и в разобранном виде они обеспечивают высокую степень устойчивости и могут выдерживать большие нагрузки. Чаще всего материал изготовления – натуральное дерево, а направляющие – из металла.
 Обратите внимание, что старые советские столы, которые умели раскладывать, имели именно такой каркас.
Обратите внимание, что старые советские столы, которые умели раскладывать, имели именно такой каркас.Недостатки Такие столы можно назвать следующими пунктами:
- Массивный внешний вид. В зависимости от стиля интерьера это может быть как преимуществом, так и недостатком. Ведь воздушной и невесомой такую модель не назовешь;
- Неудобство, связанное с тем, что в большинстве случаев сидя вы будете упираться ногами в царгу.
Раздвижной стол с выдвижным ящиком и поворотным центром
Этот тип конструкции отличается от предыдущего только конфигурацией средней выдвижной части и способом ее установки. Модели с поворотной центральной частью считается более современным и функциональным . Это объясняется тем, что увеличение обеденной зоны обеспечивается двумя вставками по центру, которые спрятаны в подстолье. Чтобы разложить такой механизм, необходимо:
Это объясняется тем, что увеличение обеденной зоны обеспечивается двумя вставками по центру, которые спрятаны в подстолье. Чтобы разложить такой механизм, необходимо:
- Раздвинуть стороны в стороны, потянув за края;
- Под столешницей вы увидите две поворотные вставки;
- Их нужно повернуть на 180 градусов, разложить и сдвинуть к ним стороны.
После трансформации в центре стола останется несколько стыковочных швов. Полезная площадь поверхности таким простым способом может увеличиться на 30-40% от первоначальной длины. По поводу сложности обращения с такой конструкцией можно сказать, что поворотный механизм удобнее выдвижного.
Двойной стол-раскладушка
он же простейший в обращении и примитивном исполнении механизм раскладки. В сложенном виде это обычный маленький столик без видимых стыков на столешнице, так как он двойной. Таким образом, ощущение такое, что толщина столешницы очень большая. Разбирается такая конструкция очень просто:
Разбирается такая конструкция очень просто:
- Необходимо снять верхнюю часть столешницы;
- Под ним вы увидите то же самое, но со швом посередине;
- Боковые стороны нужно потянуть в разные стороны;
- В середине формируется рамка и свободное пространство, размер которого точно равен первому слою. Здесь нужно вернуть верхнюю часть таблицы, снятую до этого.
Преимущество такой конструкции заключается в увеличении площади ровно вдвое по сравнению с первоначальным размером. Однако есть и недостатки:
- В зависимости от материала изготовления и размеров столешница, которую необходимо полностью снять для раскладки, может иметь внушительный вес. Поэтому может потребоваться помощь второго человека;
- В разложенном виде края стола практически не имеют опоры или опоры. Это делает конструкцию менее надежной и вводит ограничения на размещение предметов на столе. Тяжелые рекомендуется ставить ближе к краю.
Раздвижной стол с подвижными ножками
она же совершенно новый механизм который кардинально отличается от всего вышеперечисленного. Если в предыдущих случаях в процессе трансформации участвовала только столешница, то теперь в раскладке принимаю участие и я. Существует двух модификаций с аналогичным механизмом — раздвижной и складной. В сложенном виде стол ничем не отличается от обычного. Чтобы разложить раздвижную конструкцию нужно:
Если в предыдущих случаях в процессе трансформации участвовала только столешница, то теперь в раскладке принимаю участие и я. Существует двух модификаций с аналогичным механизмом — раздвижной и складной. В сложенном виде стол ничем не отличается от обычного. Чтобы разложить раздвижную конструкцию нужно:
- Держите царгу снизу и тяните на себя;
- Проржавеет, и вы увидите дополнительную столешницу;
- В некоторых моделях его нужно вытащить и повернуть, а в некоторых достаточно просто вытолкнуть.
При этом поверхность столешницы увеличивается на 30-50% без особых усилий.
Раскладной Модификация в сложенном виде выглядит как обычный стол с двухслойной столешницей. Механизм работает следующим образом:
- Тянуть на себя, удерживая короля;
- После того, как ножки полностью ушли, нужно разложить столешницу, как книгу;
- В разложенном виде его необходимо переместить в центр стола.

В итоге у вас получится столешница, площадь которой намного больше размера собранного стола. Иногда, для облегчения процесса раскладки, конструкция оснащается дополнительным подъемным механизмом , который помогает выдвигать столешницу с наименьшими усилиями. Такие столы уже можно отнести к конструкциям с полуавтоматической раскладкой. Однако у них есть одно существенное недостаток:
- Наименьший показатель долговечности по сравнению с другими механизмами;
- В разложенном виде сама столешница имеет не очень эстетичный вид, так как четко видна граница перехода основного элемента и дополнительного.
Безредукторный редуктор
Отличительной особенностью таких столов является отсутствие массивной опоры для столешницы. Это позволяет визуально сделать дизайн более изящным и не настолько громоздкий, что иногда просто необходим. Часто в качестве материала столешницы производители используют закаленное стекло, что делает стол еще «легче» и придает ему современный вид. Такие столики отлично дополнят интерьеры в стиле минимализм, лофт или хай-тек. Выдвижной механизм представляет собой металлические направляющие , которые монтируются под столешницу. Конечно, через прозрачное стекло их будет видно, но внешний вид изделия это не портит. Наоборот, это делает его еще более интересным.
Такие столики отлично дополнят интерьеры в стиле минимализм, лофт или хай-тек. Выдвижной механизм представляет собой металлические направляющие , которые монтируются под столешницу. Конечно, через прозрачное стекло их будет видно, но внешний вид изделия это не портит. Наоборот, это делает его еще более интересным.
Стол без стекла Стол без стекла обладает высокими эстетическими качествами и рядом с другими преимуществами:
- Полезная площадь поверхности может увеличиваться на 30-50% от исходной, а иногда и вдвое;
- Механизм очень плавный и легко ходит, отсутствие всевозможных заеданий и прочих сложностей. Это делает процесс верстки очень простым и не требует физических усилий.
Среди недостатков конструкций с отсутствием выдвижного ящика можно выделить:
- Относительно меньшая прочность. Причем это касается и самих направляющих, которые уступают направляющим из сбитого дерева, и самой столешницы, если речь идет о таком материале, как стекло;
- Если после раскладки длина столешницы увеличивается в два и более раза, необходимо использовать дополнительную опору;
- Стоимость таких столов может значительно превышать стоимость столов с выдвижным каркасом. Это напрямую зависит от того, какие аксессуары используются и какой материал был выбран для столешницы. Цена моделей от всемирно известных производителей может достигать 1000-3000 €.
 Это напрямую зависит от того, какие аксессуары используются и какой материал был выбран для столешницы. Цена моделей от всемирно известных производителей может достигать 1000-3000 €.
Это напрямую зависит от того, какие аксессуары используются и какой материал был выбран для столешницы. Цена моделей от всемирно известных производителей может достигать 1000-3000 €.Советы по столешнице
С одной стороны, разнообразие предлагаемых форм может сильно усложнить выбор, а с другой стороны, позволяет подобрать оптимальную модель именно для вашего помещения. Форма столешницы должна гармонировать с общим дизайном помещения и подбираться в зависимости от его площади. Также необходимо учитывать, сколько человек чаще всего будет сидеть за столом. Рассмотрим подробнее все возможные варианты:
- Квадрат форма. Такие столы могут иметь как достаточно компактные размеры в собранном виде, так и достаточно габаритные. Первый вариант будет оптимален для размещения на маленькой кухне. Более крупные модели органично смотрятся в просторных столовых или гостиных, совмещенных с кухней. Средние стандартные размеры квадратной столешницы позволяют одновременно разместить 4 человека в собранном виде и от 6 до 8 в разложенном. Этого более чем достаточно для небольшой семьи. К недостаткам ТО можно отнести пониженный уровень комфорта и ограничение по количеству посадочных мест. Это связано с наличием ножек с каждого угла, что дает возможность полноценно разместить только одного человека с конца стола;
- Прямоугольная столешница считается оптимальным решением и прекрасно вписывается как в современный, так и в классический стиль интерьера. Такие столы актуальны для использования как в просторных, так и в небольших помещениях. В собранном состоянии в них может поместиться до 6 человек, а в разложенном — все 12, что очень удобно для больших компаний.
- Круглые столы — оригинальный и нестандартный выбор. Они гармонично смотрятся как в небольших помещениях, так и в просторных. Их функция заключается в том, что они занимают гораздо меньше места, чем квадратные или прямоугольные при отсутствии углов. Этот факт облегчает передвижение по комнате. Столы круглой формы рекомендуются семьям с маленькими детьми. Это сведет к минимуму возможность получения травмы от случайного удара об острый угол. Стандартный диаметр круглой столешницы 90-110 см в собранном виде. В разложенном состоянии либо превращается в овал, либо увеличивает полезную площадь на 30-50%;
- Овальные столы также считаются более компактными и безопасными. По вместительности они практически не уступают прямоугольным моделям, что делает их очень популярными, и при этом занимают меньше места. Можно сказать, что такая форма является промежуточной между кругом и прямоугольником;
- Восьмиугольная форма встречается не у каждого производителя. Такое необычное решение, несмотря на наличие большого количества углов, очень вместительно и удобно. Такая форма четко определяет количество посадочных мест за столом и обеспечивает высокий уровень комфорта. А благодаря неординарному внешнему виду восьмиугольный стол может стать настоящим украшением и изюминкой интерьера.
 Этого более чем достаточно для небольшой семьи. К недостаткам ТО можно отнести пониженный уровень комфорта и ограничение по количеству посадочных мест. Это связано с наличием ножек с каждого угла, что дает возможность полноценно разместить только одного человека с конца стола;
Этого более чем достаточно для небольшой семьи. К недостаткам ТО можно отнести пониженный уровень комфорта и ограничение по количеству посадочных мест. Это связано с наличием ножек с каждого угла, что дает возможность полноценно разместить только одного человека с конца стола; Столы круглой формы рекомендуются семьям с маленькими детьми. Это сведет к минимуму возможность получения травмы от случайного удара об острый угол. Стандартный диаметр круглой столешницы 90-110 см в собранном виде. В разложенном состоянии либо превращается в овал, либо увеличивает полезную площадь на 30-50%;
Столы круглой формы рекомендуются семьям с маленькими детьми. Это сведет к минимуму возможность получения травмы от случайного удара об острый угол. Стандартный диаметр круглой столешницы 90-110 см в собранном виде. В разложенном состоянии либо превращается в овал, либо увеличивает полезную площадь на 30-50%;
Выбор материала
Как и другие предметы мебели, раздвижные столы могут быть изготовлены из различных материалов. Естественно, это сказывается на стоимости изделия, что дает возможность подобрать модель на разный бюджет. Выбор материала напрямую влияет на прочность и на долговечность, поэтому выбор должен быть осознанным и аргументированным. На сегодняшний день производители предлагают следующие варианты, которые мы перечислим в порядке возрастания стоимости:
- ДСП — относится к самым доступным материалам. Соответственно, прочность изделия из него будет наименьшей, но это не значит, что не стоит даже рассматривать такой вариант. При правильном уходе и эксплуатации стол из ДСП может прослужить не один год. Обратите внимание на толщину изделия. Лучшие модели изготавливаются из ламинированных листов толщиной не менее 16 – 18мм. Также особое внимание следует уделить качеству обработки торцов, которые чаще всего оклеиваются декоративной кромкой толщиной около 1 мм. Он защищает материал от влаги и препятствует его набуханию. Кромка может быть подобрана точно в тон листа ДСП, а может быть на несколько тонов темнее или светлее и оттенять основной цвет. Ламинированная поверхность материала может быть самых разных тонов, имитировать поверхность натурального дерева и иметь соответствующую фактуру, повторять цвет натурального камня и других материалов;
- МДФ — более прочный материал, чем ДСП за счет использования мелких фракций при производстве щепы и спекания листов под высоким давлением. МДФ относится к влагостойким материалам, что значительно облегчает уход за готовым изделием. По прочности и стоимости он превосходит ДСП, но и уступает натуральному дереву. Поверхность может иметь самые разные фактуры и цвета, поэтому модели столов из МДФ отличаются эстетичным внешним видом;
- Пластик — Очень удобный материал для изготовления обеденных слов. Он не боится влаги, что позволяет протирать изделие влажной губкой или накрывать столы возле раковины. Дизайнеры часто предпочитают этот материал остальным из-за его гибкости и возможности делать предметы самых невообразимых форм. Пластиковые столы станут прекрасным дополнением к современным интерьерам, например, в стиле хай-тек. Несмотря на свою легкость, конструкции обладают достаточной прочностью и ударопрочностью. Единственное, чего не любит пластик, так это высоких температур и механических воздействий. В связи с этим не рекомендуется удалять грязь с поверхности с помощью жестких губок, ставить горячую посуду без подставок или накрывать столы возле печей;
- Натуральное дерево — Этот материал всегда считался одним из лучших для изготовления мебели, и столы не исключение. Натуральная древесина имеет множество достоинств – повышенная прочность, долговечность, надежность, природная красота. Деревянные изделия не нуждаются в дополнительной декоративной отделке; они уже выглядят дорого и презентабельно. Кроме того, это экологически чистый материал, не вызывающий аллергических реакций. Древесина может использоваться как для изготовления всех элементов конструкции, так и отдельно для каркаса или столешницы. Обязательно убедитесь, что поверхность хорошо отшлифована, пропитана специальным раствором и покрыта лаком;
- Стекло в основном используется для изготовления столешниц. Очень редко, но все же можно встретить модели полностью из стекла. Выглядят они действительно эффектно, но и стоимость их находится на соответствующем уровне. Самая распространенная комбинация – стеклянная столешница + металлическая или деревянная рама. При изготовлении используется не обычное стекло, а более толстое, закаленное. Это практически исключает возможность появления мелких царапин. Но если вы ударитесь о торец столешницы, то вполне можете отколоть небольшой осколок. Стекло не боится перепадов температуры, поэтому на его поверхность без последствий можно ставить горячую посуду. При бережном обращении стеклянные изделия могут прослужить долгие годы и не потерять своей первоначальной привлекательности;
- Металл — прочный и надежный материал, из которого в большинстве случаев делают каркас и направляющие. Чаще всего используется алюминий или сталь, что значительно облегчает вес всей конструкции. Также повышается устойчивость изделия к воздействию негативных внешних факторов – коррозии, влаге, плесени, впитыванию неприятных запахов. Столы полностью из металла встречаются редко. Они смотрятся уместно только в ультрасовременных интерьерах. Чаще всего столешницы делают из других материалов – стекла, дерева, камня;
- Натуральный или искусственный бриллиант В основном используется в декоративных целях для отделки или изготовления столешниц. Такие столы очень тяжелые и дорогие. Искусственный камень легче и не такой пористый, как натуральный. Но в обоих случаях следует избегать ударов о поверхность. Это приведет к образованию трещин и сколов.
 Он защищает материал от влаги и препятствует его набуханию. Кромка может быть подобрана точно в тон листа ДСП, а может быть на несколько тонов темнее или светлее и оттенять основной цвет. Ламинированная поверхность материала может быть самых разных тонов, имитировать поверхность натурального дерева и иметь соответствующую фактуру, повторять цвет натурального камня и других материалов;
Он защищает материал от влаги и препятствует его набуханию. Кромка может быть подобрана точно в тон листа ДСП, а может быть на несколько тонов темнее или светлее и оттенять основной цвет. Ламинированная поверхность материала может быть самых разных тонов, имитировать поверхность натурального дерева и иметь соответствующую фактуру, повторять цвет натурального камня и других материалов; Дизайнеры часто предпочитают этот материал остальным из-за его гибкости и возможности делать предметы самых невообразимых форм. Пластиковые столы станут прекрасным дополнением к современным интерьерам, например, в стиле хай-тек. Несмотря на свою легкость, конструкции обладают достаточной прочностью и ударопрочностью. Единственное, чего не любит пластик, так это высоких температур и механических воздействий. В связи с этим не рекомендуется удалять грязь с поверхности с помощью жестких губок, ставить горячую посуду без подставок или накрывать столы возле печей;
Дизайнеры часто предпочитают этот материал остальным из-за его гибкости и возможности делать предметы самых невообразимых форм. Пластиковые столы станут прекрасным дополнением к современным интерьерам, например, в стиле хай-тек. Несмотря на свою легкость, конструкции обладают достаточной прочностью и ударопрочностью. Единственное, чего не любит пластик, так это высоких температур и механических воздействий. В связи с этим не рекомендуется удалять грязь с поверхности с помощью жестких губок, ставить горячую посуду без подставок или накрывать столы возле печей; Древесина может использоваться как для изготовления всех элементов конструкции, так и отдельно для каркаса или столешницы. Обязательно убедитесь, что поверхность хорошо отшлифована, пропитана специальным раствором и покрыта лаком;
Древесина может использоваться как для изготовления всех элементов конструкции, так и отдельно для каркаса или столешницы. Обязательно убедитесь, что поверхность хорошо отшлифована, пропитана специальным раствором и покрыта лаком; Чаще всего используется алюминий или сталь, что значительно облегчает вес всей конструкции. Также повышается устойчивость изделия к воздействию негативных внешних факторов – коррозии, влаге, плесени, впитыванию неприятных запахов. Столы полностью из металла встречаются редко. Они смотрятся уместно только в ультрасовременных интерьерах. Чаще всего столешницы делают из других материалов – стекла, дерева, камня;
Чаще всего используется алюминий или сталь, что значительно облегчает вес всей конструкции. Также повышается устойчивость изделия к воздействию негативных внешних факторов – коррозии, влаге, плесени, впитыванию неприятных запахов. Столы полностью из металла встречаются редко. Они смотрятся уместно только в ультрасовременных интерьерах. Чаще всего столешницы делают из других материалов – стекла, дерева, камня;Особенности выбора фурнитуры
От качества фурнитуры будет зависеть долговечность Не только раздвижных столов, но и удобство их эксплуатации. Качественные направляющие способны обеспечить легкое и плавное движение механизма, отсутствие заеданий, люфтов и перекосов, которые могут привести к преждевременному выходу из строя. В случае экономии, и приобретения более доступной фурнитуры, вы рискуете ежегодно тратить дополнительные средства на ремонт отдельных элементов. Поэтому обратите внимание на следующие моменты:
В случае экономии, и приобретения более доступной фурнитуры, вы рискуете ежегодно тратить дополнительные средства на ремонт отдельных элементов. Поэтому обратите внимание на следующие моменты:
- Материал направляющих должен быть надежным и прочным. Лучше, если это будет металл в сочетании с пластиком;
- Обязательно попробуйте механизм в действии. Следите за тем, чтобы процесс верстки не требовал чрезмерных физических усилий;
- Производители, гарантирующие заводское качество своей продукции, часто маркируют ее соответствующим образом. Это может быть комбинация букв и цифр, номер магазина или логотип компании. Внимательно осмотрите отдельные элементы на наличие похожих символов;
Одним словом, метизы – это тот элемент раздвижных конструкций, на котором не рекомендуется экономить.
Как работать с раздвижными столами
Каждая модель раздвижного стола обязательна будет иметь инструкцию где указаны правила эксплуатации. Соблюдение их продлит срок службы изделия. В зависимости от материала изготовления есть и общие рекомендации:
Соблюдение их продлит срок службы изделия. В зависимости от материала изготовления есть и общие рекомендации:
- Столы деревянные протирать специальными полиролями и только мягкой тканью. Запрещается использовать агрессивные чистящие средства, губки из твердого металла, подвергать поверхность длительному воздействию влаги;
- Стекло столов требует использования специальных моющих средств, которые указаны в инструкции. Торцевые удары и контакт с посудой, только что снятой с огня, запрещены;
- Столешницы из натурального или искусственного камня должны быть защищены от ударов. Протирайте поверхность только мягкой тканью. Не используйте жесткие скребки для удаления пятен.
При соблюдении всех требований раздвижные столы могут оправдать свою высокую стоимость долгим сроком службы и отличным внешним видом.
Метки:Таблица
Библиотека изображений слайдов | Невинс
Круглый верх
X База
Квадратный верх
X База
Прямоугольная верхняя часть
X Основание
Верх 9 листьев1101 X База
Круглый верх
4-столбцовая основа X
Квадратный верх
4-столбцовая основа X
Прямоугольная верхняя часть
T Основание
Круглый верх
X База
Квадратный верх
X База
Четверть круглая верхняя часть
X База
Верхняя часть моста
X База
Круглый верх
4-столбцовая основа X
Квадратный верх
4-столбцовая основа X
Прямоугольная верхняя часть
T Основание
Трапеция Топ
Т База
Полукруглый верх
T База
Crescent Top
T База
Верхняя часть моста
T Основание
Четверть раунда Топ
Т База
Арочный прямоугольный верх
T Основание
Презентация Top
T Base
Прямоугольная верхняя часть
C Основание
Трапециевидная верхняя часть
C Основание
Полукруглый верх
C База
Crescent Top
C База
Верхняя часть моста
C Основание
Четверть круглая верхняя часть
C Основание
Арочный прямоугольный верх
C Основание
Презентация Top
C Base
Круглый Топ
X База
Квадратный верх
X База
Круглый верх
4-столбцовая основа X
Квадратный верх
4-столбцовая основа X
Прямоугольная верхняя часть
2-колонная Т-образная основа
Верх для гоночной трассы
2-колонная Т-образная база
Изогнутая верхняя часть
2-колонная Т-образная база
Boatshape Top
2-колонная Т-образная база
Овальная круглая вершина
2-колонная Т-образная база
Клин Топ
Прямоугольная верхняя часть
Половина топа в форме лодочки
D-образный топ
Топ на танкетке с подъемником для телевизора
Прямоугольная столешница с подъемником для телевизора
Половина топа в форме лодочки с подъемником для телевизора
Верхняя часть D-образной формы с подъемником для телевизора
Топ на танкетке без TV Lift
Прямоугольная столешница без подъемника для телевизора
Половина топа в форме лодочки без подъемника для телевизора
Верхняя часть D-образной формы без подъемника для телевизора
Круглый верх
X База
Круглый верх
4-столбцовая основа X
Квадратный верх
X База
Квадратный верх
4-столбцовая основа X
Овальная вершина
4-столбцовая основа X
Штифт регулируемый
Штифт регулируемый
Пневматический регулируемый
Переворачивание
Переворачивание механизма Нестера
Переворачивание механизма Нестера
Переворачивание механизма Нестера
Переворачивание
Механизм Genu-Flip
Переворачивание
Механизм Genu-Flip, панель Modesty
Складные топы
60-96″L
Складные топы
60-96″L
Складной
42-48″Д
C1 Кастер
C1 Кастер
C5 Кастер
C5 Кастер
Ламинат или шпон Фиксированный
Переворот/фальцовка ламината/шпона
Ткань
KrystalCast®
Перфорированный металл
Модель #MPM2
Управление проводами
Модель #MPM2
Перфорированный металл
Модель #MPM4
Арочный прямоугольник
Арочный прямоугольник
Презентация | Арочные прямоугольники
Прямоугольники | Арочные прямоугольники
Прямоугольники | Арочные прямоугольники
Полумесяцы
Прямоугольники | Полукруглые
Прямоугольники | Презентация
Прямоугольники | Четверть раунда
Прямоугольники
Прямоугольники
Трапеции
от 72″Д до 96″Д
от 120 до 144 дюймов
от 168 до 216 дюймов
240 дюймов
288″Д
Cove Round
3 Доступны конфигурации разъема/питания (доступны дополнительные варианты отделки)
Cove
Доступны 8 конфигураций разъема/питания (доступны дополнительные варианты отделки)
Mini Oasis
3 Доступны конфигурации разъема / питания (доступны дополнительные варианты отделки)
Вилла
4 Доступны конфигурации разъема / питания (доступны дополнительные варианты отделки)
Interact G2
Стандартная конфигурация разъема включает питание и данные. Доступны альтернативные разъемы. (Доступны дополнительные варианты отделки)
Доступны альтернативные разъемы. (Доступны дополнительные варианты отделки)
Интерфейс G2
Стандартная конфигурация разъема включает питание и данные. Доступны альтернативные разъемы. (Доступны дополнительные варианты отделки)
Стандартная конфигурация разъема Viper
включает питание и данные. Доступны альтернативные разъемы. (Доступны дополнительные варианты отделки)
Marina
2 стандартных конфигурации, с технологией совместного использования мультимедиа или без нее (доступны дополнительные варианты отделки)
Стандартная конфигурация разъема Elf
включает питание и данные. Доступны альтернативные разъемы.
Доступны альтернативные разъемы.
Убежище
6 Доступны конфигурации разъема/питания (доступны дополнительные варианты отделки)
Metal Legway
Простая установка с защелкой позволяет многократно менять положение крышки.
Пластиковая ножка
Открывается/закрывается защелками и крепится к колонне с помощью высокопрочной ленты.
Зажимы для крепления проводов
Гибкий пластиковый зажим.
Дверь для доступа к проводам
Подпружиненная, самозакрывающаяся стальная дверь.